It seems we can’t find what you’re looking for. Perhaps searching can help.
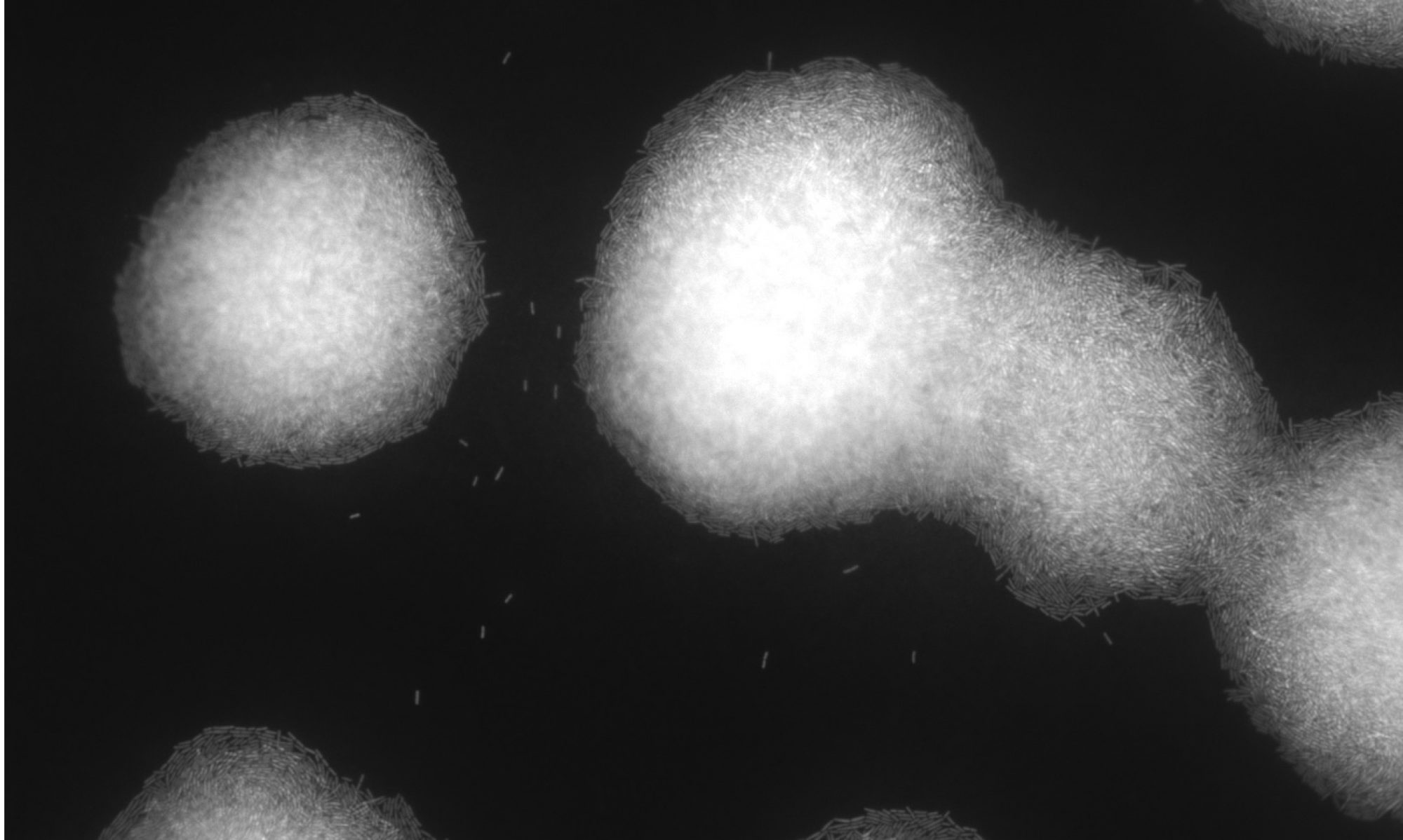

Open Kitchen Science on metabolism of vaginal microbes
Open Kitchen Science on metabolism of vaginal microbes
It seems we can’t find what you’re looking for. Perhaps searching can help.