We have finally made a start with our Citizen science project! The goal in this 2 year project is to isolate specific vaginal lactic acid bacteria (so-called ‘crispatus‘ bacteria) with a group of citizen scientists. These bacteria are associated with favorable vaginal health. We want to design a product with a mixture of these special crispatus strains and do research because we believe they could have potential health benefits, for example helping to prevent urinary tract infections or other vaginal infections. The citizen scientists/donors have a say about the destination of the bacteria. For more information find our website (in Dutch) and the public lecture (in Dutch).
This project is being carried out in collaboration with Vrije Universiteit Amsterdam, the organic femcare company Yoni, microbial genetics experts of Baseclear and Winclove probiotics. The crispatus foundation will take care of the strain collection and represent the citizen scientists. In 2022 we received a subsidy from the ZonMW / HealthHolland Citizen science for Healthcare fund with a total amount of 220,000 euros! In addition, we received almost 15,000 euros from the Voor Vrouwen Door Vrouwen fund to organize the research not only in the labs of the university, but also to get it ‘on the road’. And on her birthday, Daniela, one of our citizen scientists, raised an additional amount to purchase the Nanopore sequencing kits that we so wanted to add to our molecular toolbox. This will soon enable us to easily read out DNA in the lab. A good starting position!
We are hard at work in the lab preparing this practical and we were very lucky to hire a great analist, Shardelice Illidge, who is speeding up the experiments. We prepare all methods so that they work well in the practical. My own modest employment also started on November 1 (1 day a week), which means I can spend even more time on this project.
One cool tool we would like to present here: a colorimetric LAMP test for quick and easy identifaction of crispatus colonies from an agarose plate. We have added the protocol and some control experiments . We will use it so that citizen scientists will be able to select their own crispatus strains grown from a vaginal swab on a TSB-serum plate. Find the protocol on FigShare and on the protocols page. I will add a file with the accompanying data in a later stage. We will now prepare for submitting for peer review.
Results from LAMP test for Lactobacillus crispatus identification with NC = negative control PC = positive control (genomic DNA from L. crispatus RL100, pl = L. plantarum cr = L. crispatus bul = L. delbrueckii subsp. bulgaricus and gas = L. gasseri
On the Gardnerella front: we have finalized a manuscript about glycogen growth and amylase activity of Gardnerella species and amylase activity in vaginal samples. Although we are very proud of the work and believe it is an important addition to the field, it is also clear that we do not fully understand where the differences between the various species are coming from. All of them carry a copy of the previously identified amylopullulanase (here and here) and still some have very high activity (swidsinski and leopoldii) whereas other very low (piotii). We are very lucky to have Sara Morselli joining are lab to further figure this out. She is a PhD student from Bologna (Antonella Morongoni lab).
Find the preprint here or download here [PDF]. Unfortunately the BiorXiv server does not allow upload of the video that is a supplementary file of the manuscript so please find it here.
Puck Achterberg has started her PhD studies with a very exciting ecological approach to understanding the principles driving a Lactobacillus dominated vaginal microbiome. She will be working at the TU Delft in the lab of Professor Mark van Loosdrecht in the Environmental Biotechnology section but will also spend periods in Amsterdam detailing the metabolic requirements of Lactobacillus crispatus.
Other news: together with Jeanine Roeters-Lennep and Monique Mulders from the ErasmusMC we applied and unfortunately got rejected for a reprogrants. We would like to share our proposal here. We aim to study utilization of host lipids by vaginal bacteria after some fascinating preliminary results. We will be carrying out some initial experiments to get started on the project. Will keep you updated!
Since the start in 2017, the research presented on this blog has been unfunded, and made possible by the hospitability of the Vije Universiteit Amsterdam. But things have been changing. The exciting research findings of the past four years have made us more hungry and more ambitious. Also, the focus has shifted from a pure open science ambition towards a more inclusive science. Breaking down barriers between the university and the public: citizen science! (of course continuing as much as possible with the principles for Open Kitchen Science)
To this end, we started a foundation to strengthen this aim together with Wilbert Sybesma and Remco Kort. It was beautifully designed by Michelle Haak. https://crispatus.org/
When ZonMW (Netherlands Organization for Health Research and Development) announced the Citizen Science for Health and Care (CS4GZ) program I was immediately excited. This would give us a chance to finally involve ‘normal’ interested women to learn and study their own vaginal physiology. But more importantly it would allow us to move forward with our plans to develop a probiotic to modify the vaginal microbiome. We wanted to work with a group of donors who were aware and involved, who had ownership of the product and the research and development process in all stages: citizen scientists!
A first workshop was held in September at Micropia, facilitated by a workshop-voucher by ZonMW. Find the article on their website (Dutch) https://www.zonmw.nl/nl/actueel/nieuws/detail/item/workshopvouchers-zo-besteed-je-ze/?utm_source=linkedin&utm_medium=nieuws&utm_campaign=lsh
Since in this project we will work with a larger group of industrial partners, researchers and citizen scientists, unfortunately we will not be able to share all plans, agreements, contracts and datasets that belong to the project. But of course we will continue sharing as much as possible, also on REBLAB.
On November 9th we will apply with a consortium of partners, including several consortium partners. Fingers crossed!!!
Lots of progress at the Reblab front! I have uploaded several datasets/slides and reports to Figshare and will give s hort introduction here:
First, we have a very interesting finding around Gardnerella amylase activity. Leon Steenbergen has finalized his masters thesis this summer and I presented some of his most important findings and some of our own in a presentation during the LAB symposium. Find the slides here.
Next, I finally managed to finalize the first retrospective data analysis together with the Helius study cohort which unfortunately leaded to a negative result. Previously I have shared the research proposal. Below find the FigShare link to the results report (negative results) + statistics (thanks Douwe Molenaar)! and of course thanks to the Helius group including Henrike van Galenkamp – van der Ploeg, Marieke Snijder, Maria Prins and Koos Zwinderman.
Happy new year everyone! To start this year off I would like to share the work of Alexander Woudstra, a 2nd year master student at the Vrije Universiteit Amsterdam. As part of his internship he has studied glycogen metabolism of the vaginal bacterial species Lactobacillus crispatus and Gardnerella vaginalis. This host-derived glycan is the most abundant vaginal carbohydrate source and thought of as vital as a acarbon and energy source.
Previously, Deborah Jekel and Ritesh Panchoe carried out growth experiments with L. crispatus and G.vaginalis on glucose and glycogen. They noticed that in the presence of glucose, L. crispatus does not break down glycogen . Furthermore, a putative pullulanase type I gene was identified in L. crispatus strains that is involved in breakdown of glycogen. Lastly, it was shown that G. vaginalis has a longer lag phase when grown on either glucose or glycogen.
Alexander further studied growth of these species on glycogen and glucose, looking specifically at lag phase, and preferred carbon source when presented with both glucose and glycogen. He also studied inhibition of growth by the alpha-glycosidase inhibitor acarbose, which inhibited growth on glycogen of both Gardnerella and Lactobacillus but also – surprisingly- showed inhibiton of Gardnerella growth on glucose.
Find his full thesis here. We wish Alexander all the best on his future steps!
I have recently been reading about the role of the vaginal microbiota and fertility. Multiple reports have now emerged that the microbiome affects succes rates in fertility treatment. Women colonized by lactobacilli seem to have higher success rates when using artificial reproductive techniques such as in vitro fertilization treatment.
Next question is of course: How does this work? What is the mechanism?
There are lots of possible answers coming from immunological, endocrine and gynaecological perspective that will all have some grain of truth. The vaginal microbiome may be a secondary outcome of a host endocrine factor which may result both in lowered Lactobacillus colonization as well as reduced fertility.
I hypothesize that there may be a metabolic mechanism too in which the bacterial members of the dysbiotic microbiome can contribute autonomously to subfertility. Specifically, colonization of high numbers of Gardnerellavaginalis, a glycogen-degrading species, may render the endometrium less conceptive to embryo transfer due to bacterial breakdown of endometrial glycogen stores.
There are a few phenomena on which this hypothesis is based, that I will outline using the respective literature:
Firstly: our own research shows that Gardnerellavaginalis can breakdown glycogen efficiently. It will most likely secrete its enzymes to access external glycogen.
Secondly: This is one of several older papers demonstrating an association between subfertility and lowered endometrial glycogen levels.
This recent review by Matthew Dean stresses the importance of these endometrial glycogen stores for implantation (although most research is done in rodents).
Lastly, at the Keystone conference in Cape Town, Caroline Mitchell reminded us all of the open access between uterus and vagina in women who are not pregnant. Many bigger particles can freely diffuse from the vagina into the uterus, so why would the glycogen-degrading enzymes of Gardnerella not access endometrium glycogen?
However, there is one big phenomenon opposing this hypothesis. Women colonized by Gardnerella do not generally show subfertility (as fas as I am aware), so it may be true only in certain conditions or only in the specific case of artificial reproductive techniques. (or it may not be true at all…)
In short, I am very curious to look at the link between endometrial glycogen and the vaginal microbiome. Could bacterial enzymes reach endometrial glycogen stores and render the endometrium less suitable for embryo transfer or implantation?
Ritesh Panchoe and Rosanne Hertzberger, REBLAB / Group of Prof. Dr. Remco Kort, Systems Biology Lab, AIMMS, Vrije Universiteit Amsterdam, O|2 location De Boelelaan 1085, NL-1081HV Amsterdam, The Netherlands
Find
the corresponding data file here…
Summary of the finding
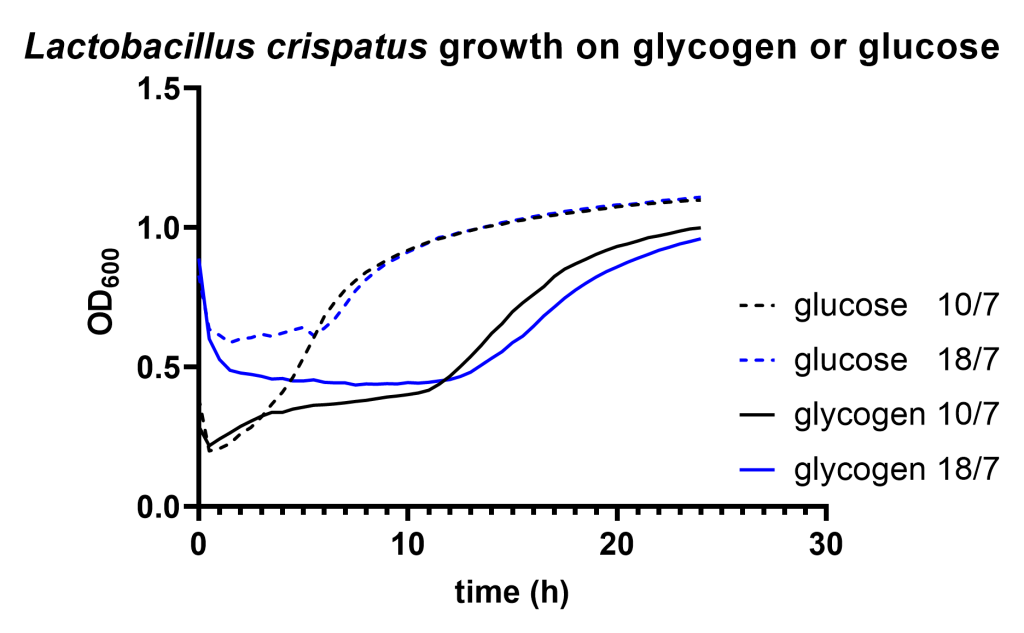
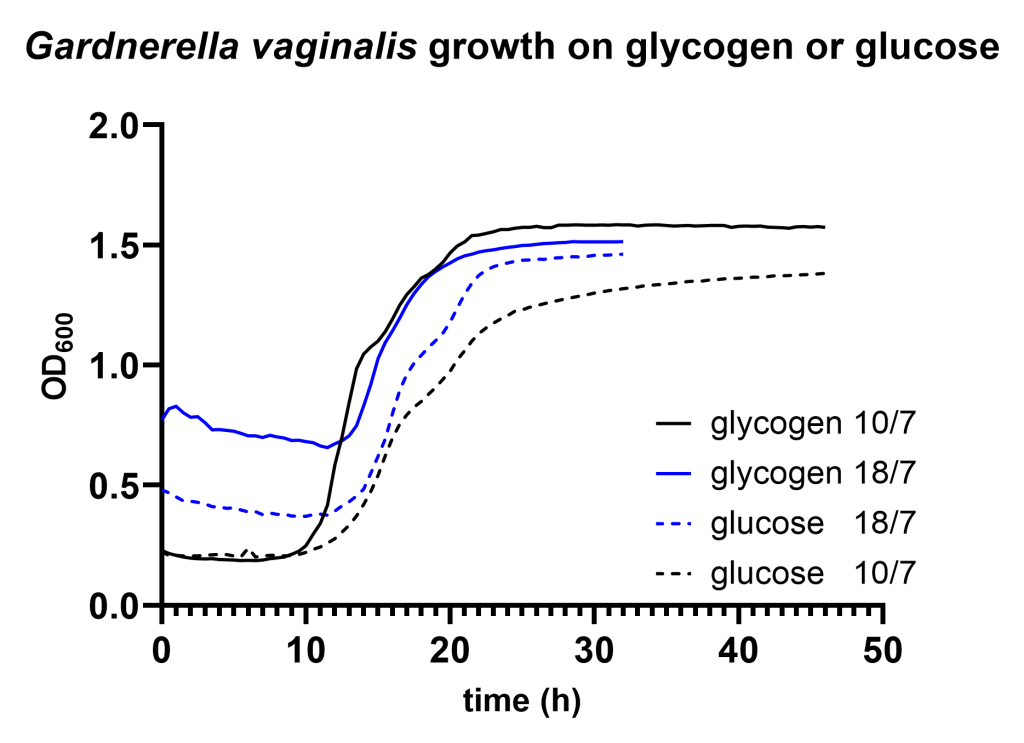
After a pregrowth step on glucose, Lactobacillus crispatus strain RL_010 shows a significant lag phase on glycogen compared to glucose. This is consistent with the differential regulation of the glycogen degradation machinery which we reported before. Gardnerella vaginalis does not show such a difference in lag phase but does exhibit a long pause before starting growth on either glucose or glycogen.
Introduction
Previously we focused on growth of various strains of Lactobacillus crispatus on glycogen and glucose. We identified the role of a type 1 pullulanase, and we observed how glycogen breakdown ability is downregulated in the presence of glucose. Since a few months we have our anaerobic chamber up and running and we are now able to include Gardnerella vaginalis in our growth studies. Gardnerella belongs to the family Bifidobacteriaceae and is one of the prominent members of the dysbiotic vaginal community.
By studying glycogen metabolism in members of both the Lactobacillus-dominated and dysbiotic community we try to understand the fluctuations in vaginal glycogen content in reproductive-age women from a microbial perspective. In general, it is hypothesized that Lactobacillus utilizes glycogen as its primary carbon source to acidify the vagina. However, in the absence of a dominant Lactobacillus in the vaginal community (‘dysbiotic state’ or community state type IV) vaginal glycogen is reduced. We hypothesize that members of the dysbiotic vaginal community may be responsible for this enhanced glycogen degradation. The strong increase in absolute numbers of bacteria in the dysbiotic state increases carbon and energy requirements of the vaginal microbial community. During Ritesh’ internship we analyzed growth of Gardnerella vaginalis and Lactobacillus crispatus on glycogen to growth on glucose.
Methods
Gardnerella vaginalisDSM 4944 and Lactobacillus crispatus RL_10 were pregrown in liquid NYCIII medium [PDF] with 0,5% glucose and incubated anaerobically for 48 hours in an anaerobic chamber filled with a gas mix of 95% N2 and 5% CO2. Find full inoculation details here [PDF], in these experiments we inoculated anaerobically.
This preculture was tenfold diluted in NYCIII medium containing either 0,5% glucose, 0,5% glycogen or water as a control in flat-bottom transparent 96 well plate (Greiner 655161). The volume in each well was 380 uL to leave as little headspace as possible. The plates were sealed-off with adhesive film (VWR 60941-072).
This experiment should not be considered as anaerobic. Although the 96-well plate was prepared in the anaerobic chamber, it was subsequently taken to the plate reader outside of the chamber. It is safe to assume that part of this growth experiment should be considered aerobic.
We performed this experiment on three separate occasions and show the results of two individual experiments in the graph.
Results
We find that Lactobacillus crispatus shows a considerable longer lag phase on glycogen compared to glucose after it has been pregrown on glucose (Figure 1A). Although growth of Gardnerella vaginalis resulted in a very long lag phase (>10 hours) on both carbon sources, we did not detect any difference in lag phase between growth on glycogen compared to glucose (Figure 1B).
Figure 1A: optical density of Lactobacillus crispatus strain RL_010 growing on NYCIII medium supplemented with 5 mg/mL glucose (dashed lines) or 5 mg/mL glycogen (continuous lines) of two different experiments on two different dates. The dates are indicated in the legend, find the corresponding data files here.Figure 1B: optical density of Gardnerella vaginalis growing on NYCIII medium supplemented with 5 mg/mL glucose (dashed lines) or 5 mg/mL glycogen (continuous lines) of two different experiments on two different dates. The dates are indicated in the legend, find the corresponding data files here.
The lag phase of Lactobacillus crispatus may be explained by the repressed expression of the type 1 pullulanase in the presence of glucose. There are several other findings that support carbon catabolite regulation of this gene associated with glycogen metabolism: 1) previously, we were unable to detect glycogen degradation activity in L. crispatus pellets after growth on glucose. 2) The type 1 pullulanase gene of L. crispatus has a palindromic region upstream of its start site showing similarities to carbon responsive elements in Lactobacillus plantarumhttps://www.ncbi.nlm.nih.gov/pmc/articles/PMC5986886/. 3) The expression of the ortholog pullulanase gene in L. acidophilus is also repressed by glucose. None of these findings provide conclusive evidence but they are all in line with repressed expression of the type 1 pullulanase in presence of glucose.
The ~10 hours lag phase
of Gardnerella
vaginalis on glycogen is likely due to other circumstances, for
instance oxidative stress due to influx of oxygen through the microplate seal. The
results do not exclude that Gardnerella regulates its genes involved in glycogen
degradation, but under these experimental conditions Gardnerella grows
as readily on glycogen as on glucose.
-Warning, wild speculation ahead- This difference in regulation of the glycogen degradation machinery of these bacterial species, that are associated with distinct microbial signatures of the vagina, may have implications on the bacterial dynamics in the human vagina. If a L. crispatus culture encounters glucose in the vagina, it is likely that it will continue growth on glucose and thereby stop expressing its enzymes needed for glycogen breakdown.
After depletion of glucose the bacteria will have to switch back to using vaginal glycogen but need some time to accomplish this metabolic switch. In contrast: if we find in further experiments that Gardnerella vaginalis constitutively expresses its glycogen degradation enzymes, these bacteria can continue growing on glycogen and may even be capable of utilizing both glycogen and glucose in paralel. It may even be beneficial for Gardnerella in bacterial competition to leave trace amounts of glucose during glycogen degradation (“chew more than you eat”), to provoke this metabolic shift in Lactobacillus crispatus.
Next
experiments
-Alexander Woudstra is currently repeating these
experiments in the anaerobic chamber. Although we do not have a plate reader in
there, we do have a 37° heating block.
-A simple experiment that would confirm the
role of glycogen in these growth dynamics is by pre-growing on glycogen. This
should eliminate the L crispatus lag-phase on glycogen.
-Lastly, Alexander is also attempting to grow these bacteria on both
glucose and glycogen. If our hypothesis is correct, Lactobacillus crispatus should
degrade glucose first and then switch to glycogen. Gardnerella vaginalis
should be capable of utilizing both substrates simultaneously.
Practical notes on the experiments:
The data file has several other experimental conditions that were included. For instance: growth of Lactobacillus iners, growth on galactose and maltose and growth of Gardnerella vaginalis supplemented with supernatants from previous growth experiments. None of these experimental conditions provided results that we would like to discuss extensively at this point.
Ritesh has repeated this experiment on one more day (July 16th), but during this experiment the controls were contaminated. Also, for some unknown reason L crispatus did not show growth on glycogen during this experiment. We have omitted these lines from the graph but the data can be found in a separate tab in the data file.
The increased OD600 measurements at the onset of some growth experiments were likely due to air bubbles under the plate seal.
Some more
practical findings from his internship:
-the anaerobic chamber is key in growing Gardnerella vaginalis and Lactobacillus iners. Previously we were attempting to grow them in anaerobic jars, but this means that the handling and pipetting is happening in the presence of oxygen and this means rapid decline of viability. In this project we used the anaerobic chamber for the first time and it worked out well.
-However,
we are still experiencing problems with reproducibility of growth of Lactobacillus
iners. We still have to find the best way to capture the exponential phase
and the optimal inoculation conditions.
-Starch
degradation was in this project much more heterogeneous as previously. We need to look
into the origins of this heterogeneity, and we think it may have something to
do with handling of the cultures. I believe that the findings from Deborah’s
internship about starch degradation are still credible and reproducible, but
the difference between experiments in this project were much larger and we do
not feel confident enough to share them.
-Due to these practical reasons we chose to only share this one observation on growth of Lactobacillus crispatus and Gardnerella vaginalis.
Thanks
to
We would
like to express our gratitude to Jurgen Haanstra for supervision, advise and
positive vibes.
Hi everyone! My name is
Ritesh Panchoe and Rosanne was so kind to offer me an internship position. While
I am here I will work on the characterization of bacterial enzymes involved in
glycogen degradation. The species I will focus on are Gardnerella vaginalis, Lactobacillus iners and Lactobacillus crispatus. That being said, I also want to mention
that I have a lot of fun doing my internship while being able to work with
Rosanne. Since my specialization is in infectious diseases, and not necessarily
in glycobiology, I learn a lot from her while working on this subject.
I am working with these bacteria since early
April and already have some findings which we want to share with you. In our
assays we use acetate buffer with varying pH (4.0-4.5-5.0) to try to resemble
the vaginal pH. We have been using α-amylase (read: my saliva) as a positive
control next to B. subtilis α-amylase and for now It looks like α-amylase
from both origins has a higher activity in pH 5.0 compared to pH 4.0.
We cultured G. vaginalis in liquid NYClll medium supplemented with glycogen or
glucose. After incubation G. vaginalis
shows something that looks like film formation in the 10 ml tubes (we incubated
roughly 96 hours at 37C). Fun fact: G.
vaginalis colonies on plates smells like potatoes!
Also, amyloglucosidase (Sigma product#
A7095-50ml) does not work anymore if it is stored at -20 ℃. While this might seem as an open door
because it is stated on the product label we should store at 2-8 ℃, we thought
an extra warning could be of use (since some enzymes can be stored at freezer
temperature).
Currently we’re trying new methods
for analysis of glycogen degradation enzymes. Ultimately, we would like to be
able to differentiate between the enzymes of various vaginal bacteria, since
this allows us to identify them in human specimens.
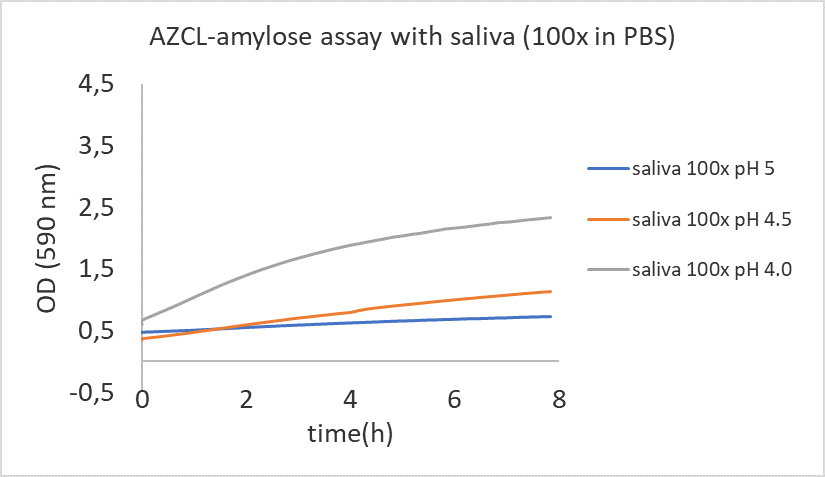
The starch-iodine assay we use only gives information about the total starch breakdown after a set incubation time. We want to be able to measure enzymatic rates over time in our samples, therefore we included the AZCL-amylose assay (protocol can be found here). We had some challenges with this assay.
Our positive controls (human saliva 100x diluted in PBS and B. subtilis amylase (60u/ml) do show amylose breakdown (see figure). We have performed this test three times, all times with the same result. But the bacterial pellets of L. crispatus give some weird results. Despite the fact that starch (which is composed of amylose and amylopectin) can be degraded, the bacterial samples included in the AZCL amylose assay showed no breakdown of amylose. This is weird.
Figure: AZCL-amylose assay with 100x saliva in PBS
We hypothesized that the xanthan gum applied
to our buffer might be the problem for the bacterial enzymes. Rosanne has added
this xantan gom as a bulk ingredient to prevent pelleting of the insoluble
granules. We tested this by omitting the xanthan gum in the buffer. As a
consequence, the AZCL-amylose granules precipitate as expected. Therefore, we
decided to do an OD measurement at 590 nm before the start of incubation. We
then incubated our samples (in a shaker at 30 ℃, we currently do not have the tools to shake at 37 ℃) and measured OD after 21
hours. There was no increase in OD values (again, controls showed correct
signals), although the positive controls did show some breakdown of the
granules. Conclusion: it is unlikely that the xanthan gum interferes with the
assay. We hypothesize now that it may have to do something with the structure
of the amylose (which is very dense).
In the coming weeks I will perform growth
experiments on G. vaginalis and L. iners, followed by HPLC of these
samples. I will follow-up as soon as I have interesting findings. I will keep
you posted!
I am Deborah Jekel and
I am a third-year bachelor student Health and Life sciences at the Vrije
Universiteit in Amsterdam. The last couple of months I have had the pleasure to work with Rosanne
and do an experimental internship. My project comprised of researching the
regulation of glycogen metabolism of Lactobacillus crispatus – a
prominent member of the vaginal microbiome. Teaser, it might possibly be carbon
catabolite repression!
Because this internship was part of an Open Kitchen Science research project, all figures, data, methods and protocols are shared. Find the entire dataset at FigShare:
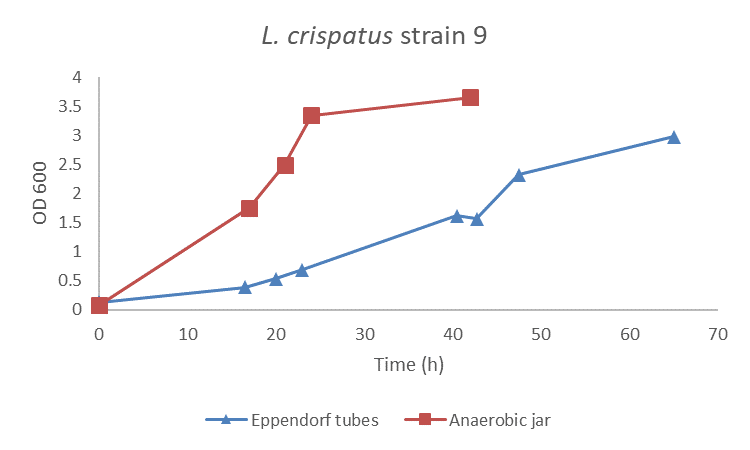
First, I spend some time on finding the optimal growth conditions for L. crispatus. I compared two methods, the use of Eppendorf tubes versus the Anaerobic jar. I cultured L. crispatus in NYCIII medium supplemented with glucose (find protocol here) and used Eppendorf tubes that I filled completely to reduce oxygen levels to a minimum and an Anaerobic jar that I pulled vacuum and filled with CO2 + N2 gas. Results presented that L. crispatus grows faster and better in the Anaerobic jar than in the Eppendorf tubes (figure 1). Of course, this is sort of an open door since we know that L. crispatus is an anaerobic bacterium, but these experiments were also good for me to get acquainted with working in the lab and to start off easy.
Figure 1: L. crispatus strain 9 grown in NYCIII medium supplemented with glucose (4.8 g/L) using an Anaerobic Jar or Eppendorf tubes. Each method was run in duplicate, a representative is shown here.
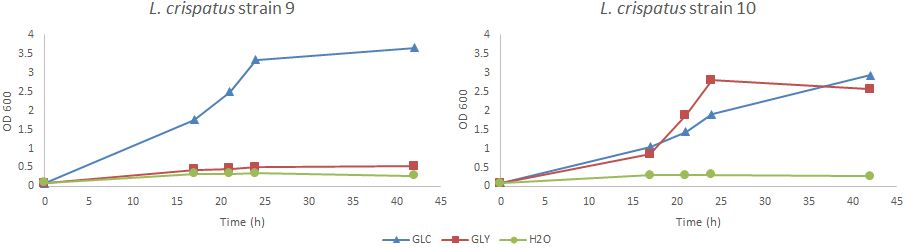
As you may know glycogen is an abundant carbohydrate in the vagina, can L. crispatus utilize glycogen to colonize the vagina? Earlier experiments completed by Rosanne already showed results that some strains can grow on glycogen-supplemented growth media (see blogpost here). I did growth experiments with L. crispatus strain 9 and 10 cultured in NYCIII medium 1.1x supplemented with glycogen, glucose or demi water (see last paragraph of NYCIII protocol). Results showed that strain 9 cannot grow on glycogen while strain 10 can, see Figure 2. This corresponds with the results of Rosanne. Could the lack of growth on glycogen by strain 9 be caused by the frameshift mutation of strain 9 in the putative type I pullulanase gene? We don’t know yet. Unfortunately, I have not been able to calculate growth rates as a result of a lack of measurements during the exponential phase of growth.
Figure 2: L. crispatus strain 9 and 10 grown in NYCIII medium supplemented with glucose (4.8 g/L), glycogen (4.8 g/L) or demi water. Experiment was run in duplicate; a representative is shown here. Abbreviations: NYC, New York City; OD, optical density; GLC, glucose; GLY, glycogen.
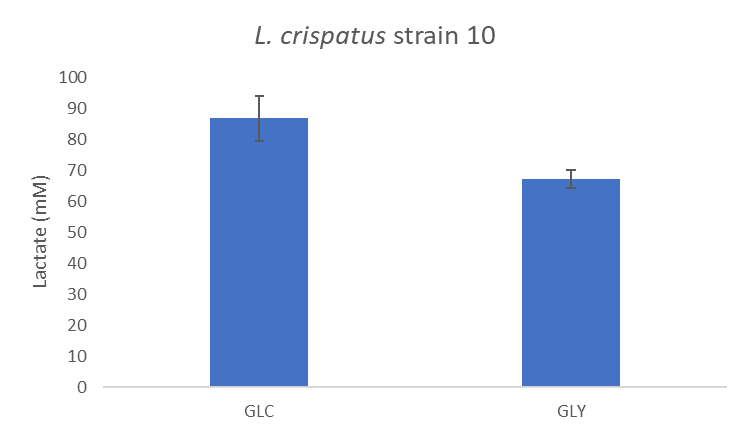
We also know that the vagina has a low pH which may be caused by lactate possibly produced by bacteria. I looked at lactate concentrations in the supernatant of L. crispatus strain 10 after growth on glucose and glycogen using HPLC (find the protocol here). We see that growth on glucose shows a higher lactate concentration than growth on glycogen, see Figure 3, but this may also be explained by the difference in cell density (optical density at 600nm) that was on average 0.4. Could it be that L. crispatus also utilizes glycogen to produce lactate and acidify the vagina?
Figure 3: Lactate concentration (mM) in supernatants of L. crispatus strains 10 after 48 hours of growth on NYCIII medium supplemented with glucose (4.8 g/L) or glycogen (4.8 g/L). Experiment was run in quintuplet; averages and standard deviations are shown.
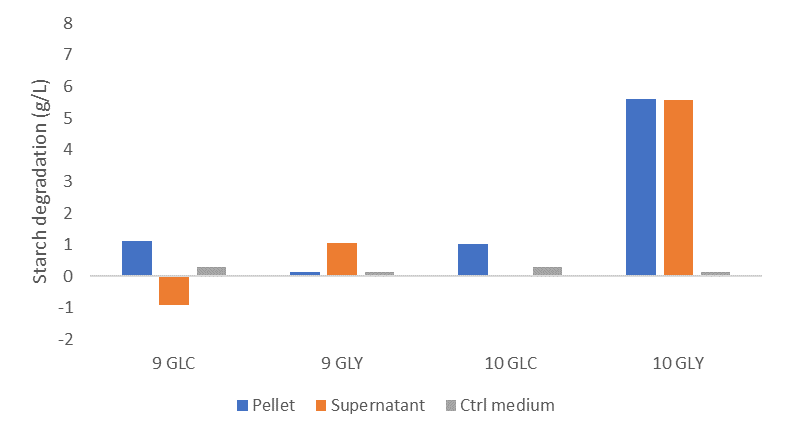
In addition, I wanted to investigate the enzymatic activity of L. crispatus. I used starch as a proxy for glycogen and carried out enzymatic assays with an iodine staining (find protocol here). I incubated the pellets and supernatants of L. crispatus strain 9 and 10 in a starch solution after growth on glycogen, glucose and demi water. I discovered that only strain 10 could degrade starch after growth on glycogen.
Figure 4: Starch degradation activity of the pellets and supernatants of L. crispatus strain 9 and 10 grown in NYCIII medium supplemented with glucose (4.8 g/L), glycogen (4.8 g/L) or demi water and after incubation in a starch solution (7.5 g/L). Experiment was run in duplicate; averages and standard deviations are shown. Abbreviations: NYC, New York City; GLC, glucose; GLY, glycogen.
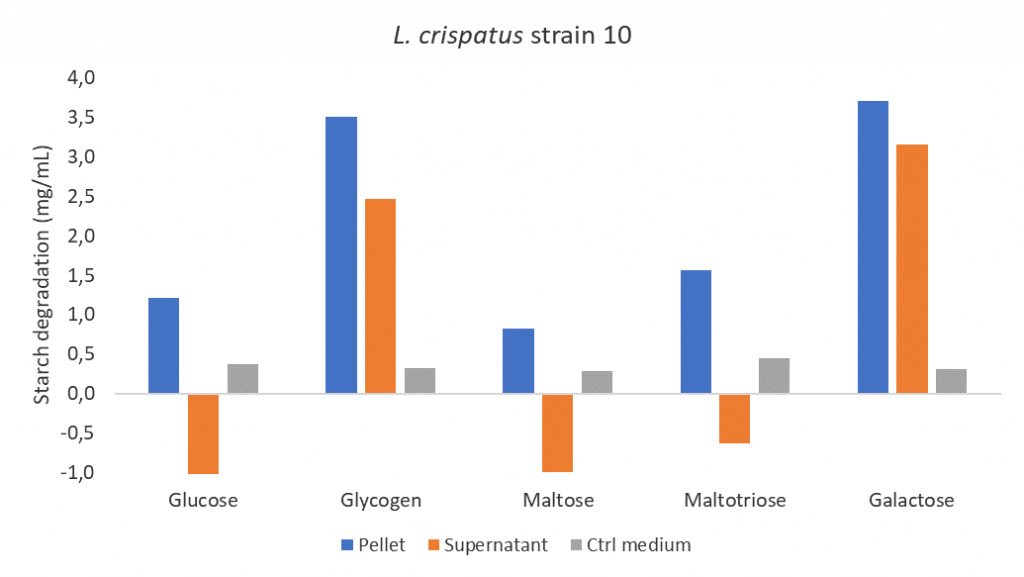
After seeing the results of the starch assay, I was curious whether growth on other carbon sources could also induce starch degradation activity. I examined maltose, maltotriose and galactose. Maltose and maltotriose are of interest because they are breakdown products of glycogen and galactose has the same structural formula as glucose but a different metabolic pathway. Interestingly, the presence of galactose during growth of L. crispatus strain 10 also induces starch degradation activity while maltose or maltotriose did not! So, some carbon sources induce starch degradation and others do not, could this be carbon catabolite repression? This would mean that L. crispatus uses the carbon sources that are most easily accessible and allow for fastest growth while inhibiting the synthesis of enzymes involved in the metabolism of secondary carbon sources. This makes sense, right? Why would L. crispatus waste energy on grinding (glycogen) when there are also mashed and grounded nutrients available (glucose, maltose or maltotriose)?
Figure 5: Starch degradation activity of the pellets and supernatants of L. crispatus strain 10 grown in NYCIII medium supplemented with glucose (4.8 g/L), glycogen (4.8 g/L), maltose (4.8 g/L), maltotriose (4.8 g/L) or galactose (4.8 g/L) and after incubation in a starch solution (7.5 g/L). Experiment was run in duplicate; averages and standard deviations are shown. Abbreviations: NYC, New York City.
That is all we have on the speculation of carbon catabolite repression in L. crispatus for now.
One more result I wanted to share: I have tried growing on two carbon sources simultaneously and performing a starch assay after 48 hours (find data here), but had some trouble interpreting the results. What I intended was to try to catch the switch from one carbon source to the other. I was hoping to find the degradation activity to correspond with the second carbon source that I added in addition to glycogen. This would prove glucose repression, instead of other regulatory mechanisms, since in the presence of glycogen and glucose the activity would remain repressed. Unfortunately, I wasn’t able to get a meaningful result, possibly because I couldn’t distinguish between growth on the different carbon sources. Experimental design needs improvement! In the future I hope we can measure glycogen concentrations and determine starch- and growth rates to be able to pinpoint exactly the moment this activity is switched on or off. Nevertheless, data of the different experiments are shown in the FigShare file and are also mentioned in my thesis (name: “double carbon sources”).
I have learned a lot these past 4 months and I want to thank Rosanne for giving me this opportunity. If you’re interested in reading my thesis, you can find it with all other data files on FigShare. Enjoy!
Hi all, I really like the format of a newsletter so I have started writing one. Find issue 1 here, and please subscribe if you want to receive these about 4x a year. You can also follow things in the lab on the REBLAB Facebook page. I will also keep on posting updates here.