
Hi there!
I am Deborah Jekel and I am a third-year bachelor student Health and Life sciences at the Vrije Universiteit in Amsterdam. The last couple of months I have had the pleasure to work with Rosanne and do an experimental internship. My project comprised of researching the regulation of glycogen metabolism of Lactobacillus crispatus – a prominent member of the vaginal microbiome. Teaser, it might possibly be carbon catabolite repression!
Because this internship was part of an Open Kitchen Science research project, all figures, data, methods and protocols are shared. Find the entire dataset at FigShare:
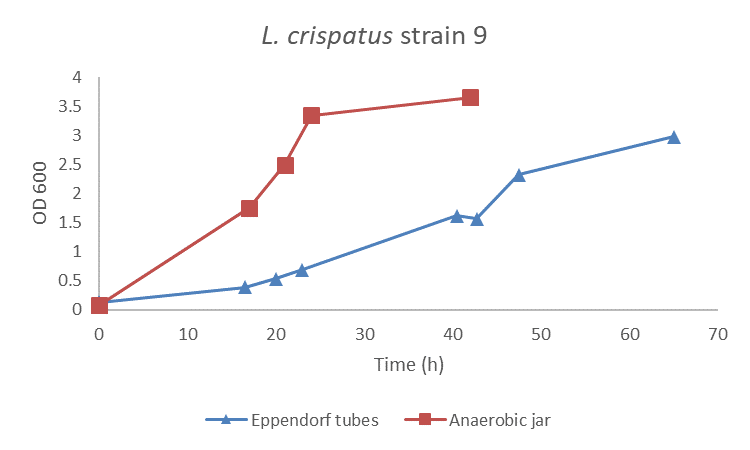
First, I spend some time on finding the optimal growth conditions for L. crispatus. I compared two methods, the use of Eppendorf tubes versus the Anaerobic jar. I cultured L. crispatus in NYCIII medium supplemented with glucose (find protocol here) and used Eppendorf tubes that I filled completely to reduce oxygen levels to a minimum and an Anaerobic jar that I pulled vacuum and filled with CO2 + N2 gas. Results presented that L. crispatus grows faster and better in the Anaerobic jar than in the Eppendorf tubes (figure 1). Of course, this is sort of an open door since we know that L. crispatus is an anaerobic bacterium, but these experiments were also good for me to get acquainted with working in the lab and to start off easy.
As you may know glycogen is an abundant carbohydrate in the vagina, can L. crispatus utilize glycogen to colonize the vagina? Earlier experiments completed by Rosanne already showed results that some strains can grow on glycogen-supplemented growth media (see blogpost here). I did growth experiments with L. crispatus strain 9 and 10 cultured in NYCIII medium 1.1x supplemented with glycogen, glucose or demi water (see last paragraph of NYCIII protocol). Results showed that strain 9 cannot grow on glycogen while strain 10 can, see Figure 2. This corresponds with the results of Rosanne. Could the lack of growth on glycogen by strain 9 be caused by the frameshift mutation of strain 9 in the putative type I pullulanase gene? We don’t know yet. Unfortunately, I have not been able to calculate growth rates as a result of a lack of measurements during the exponential phase of growth.
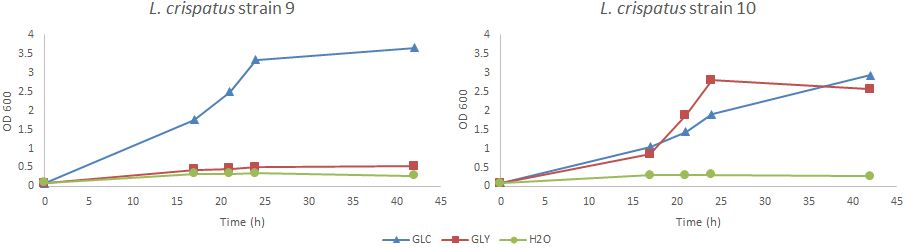
Figure 2: L. crispatus strain 9 and 10 grown in NYCIII medium supplemented with glucose (4.8 g/L), glycogen (4.8 g/L) or demi water. Experiment was run in duplicate; a representative is shown here. Abbreviations: NYC, New York City; OD, optical density; GLC, glucose; GLY, glycogen.
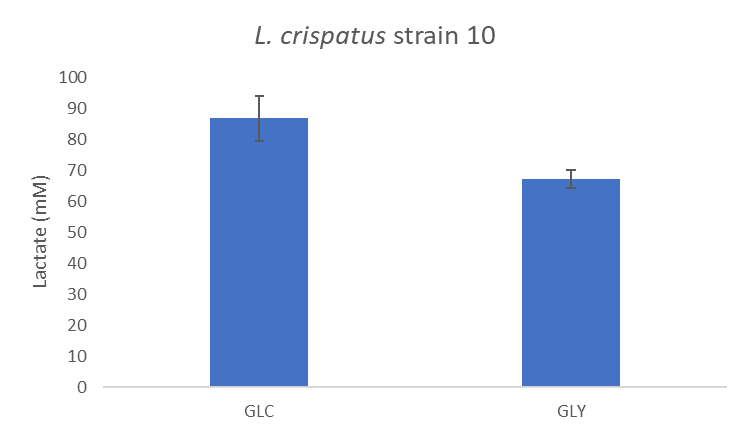
We also know that the vagina has a low pH which may be caused by lactate possibly produced by bacteria. I looked at lactate concentrations in the supernatant of L. crispatus strain 10 after growth on glucose and glycogen using HPLC (find the protocol here). We see that growth on glucose shows a higher lactate concentration than growth on glycogen, see Figure 3, but this may also be explained by the difference in cell density (optical density at 600nm) that was on average 0.4. Could it be that L. crispatus also utilizes glycogen to produce lactate and acidify the vagina?
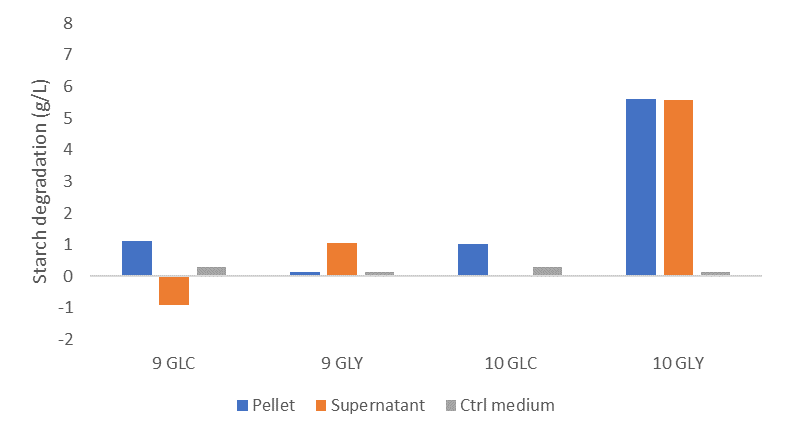
In addition, I wanted to investigate the enzymatic activity of L. crispatus. I used starch as a proxy for glycogen and carried out enzymatic assays with an iodine staining (find protocol here). I incubated the pellets and supernatants of L. crispatus strain 9 and 10 in a starch solution after growth on glycogen, glucose and demi water. I discovered that only strain 10 could degrade starch after growth on glycogen.
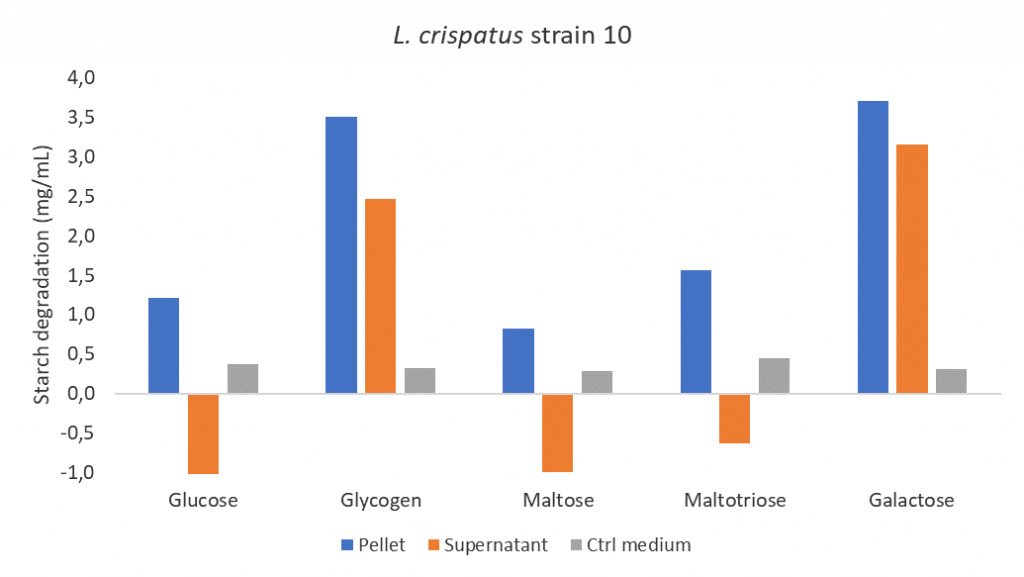
After seeing the results of the starch assay, I was curious whether growth on other carbon sources could also induce starch degradation activity. I examined maltose, maltotriose and galactose. Maltose and maltotriose are of interest because they are breakdown products of glycogen and galactose has the same structural formula as glucose but a different metabolic pathway. Interestingly, the presence of galactose during growth of L. crispatus strain 10 also induces starch degradation activity while maltose or maltotriose did not! So, some carbon sources induce starch degradation and others do not, could this be carbon catabolite repression? This would mean that L. crispatus uses the carbon sources that are most easily accessible and allow for fastest growth while inhibiting the synthesis of enzymes involved in the metabolism of secondary carbon sources. This makes sense, right? Why would L. crispatus waste energy on grinding (glycogen) when there are also mashed and grounded nutrients available (glucose, maltose or maltotriose)?
That is all we have on the speculation of carbon catabolite repression in L. crispatus for now.
One more result I wanted to share: I have tried growing on two carbon sources simultaneously and performing a starch assay after 48 hours (find data here), but had some trouble interpreting the results. What I intended was to try to catch the switch from one carbon source to the other. I was hoping to find the degradation activity to correspond with the second carbon source that I added in addition to glycogen. This would prove glucose repression, instead of other regulatory mechanisms, since in the presence of glycogen and glucose the activity would remain repressed. Unfortunately, I wasn’t able to get a meaningful result, possibly because I couldn’t distinguish between growth on the different carbon sources. Experimental design needs improvement! In the future I hope we can measure glycogen concentrations and determine starch- and growth rates to be able to pinpoint exactly the moment this activity is switched on or off. Nevertheless, data of the different experiments are shown in the FigShare file and are also mentioned in my thesis (name: “double carbon sources”).
I have learned a lot these past 4 months and I want to thank Rosanne for giving me this opportunity. If you’re interested in reading my thesis, you can find it with all other data files on FigShare. Enjoy!