
Hi all, I really like the format of a newsletter so I have started writing one. Find issue 1 here, and please subscribe if you want to receive these about 4x a year. You can also follow things in the lab on the REBLAB Facebook page. I will also keep on posting updates here.
My research
Here you can find ongoing work at REBLAB. Our most important finding so far is the connection between a type 1 pullulanase and Lactobacillus crispatus glycogen metabolism. Find the post here, and accompanying protocols and data here. Find an introductory lecture to the field and my work here. I also send out a newsletter, if you want stay up to date subscribe here
Research proposal: genetic factors of host glycogen metabolism and its relationship with the vaginal microbiome.
After visiting the Keystone conference I was wondering if genetic factors of the hostess could be related to her vaginal microbiome. Janneke van de Wijgert pointed me towards the HELIUS study, that has both SNP data and vaginal microbiome available for a few hundred participants. I wrote a proposal to compare these datasets. I hope we will be able to perform this study this year. If you are working on something comparable, let me know and perhaps we can join forces! If you have any other proposals of promising SNP’s that we should take into account in the analysis, I would be very interested to hear about this.
Summary Evidence is accumulating that glycogen released by endometrium and the cervicovaginal epithelium functions as an important carbohydrate for vaginal bacteria. Luminal glycogen is found to vary both with hormonal status and with the bacterial make-up of the reproductive tract. Here we propose to study whether host genetic factors that are involved in glycogen metabolism regulation show correlation with vaginal microbial signatures.
The multi-ethnic HELIUS cohort of Amsterdam is uniquely positioned for this analysis [1]. The vaginal microbiome of several participants was characterized previously [2], showing a variation in Lactobacillus-dominated microbiota and Lactobacillus-depleted dysbiotic states. This latter microbial state is associated with an increased risk of preterm labor and acquisition of sexually transmitted infection. Of a subset of these women (346 in total) genome wide SNP data will become available this year. We propose to analyze whether women with Lactobacillus-dominated vaginal microbiota are significantly more likely to carry certain alleles in the most prominent SNP’s of the TCF7L2 gene that are associated with type 2 diabetes. This preliminary analysis data could inform novel studies within this same HELIUS cohort to study the correlations between host vaginal glycogen synthesis, lactate concentration and vaginal microbiome.
Hypothesis: Glycogen functions as a carbon source for Lactobacillus species colonizing and acidifying the human vagina. We hypothesize that genetic changes in the glycogen synthesis pathway and its regulatory factors can affect the ability of Lactobacillus species to acidify the vagina and prevent changing to a bacterial vaginosis microbial state. Moreover, unpublished experiments have shown that bacterial members of this dysbiotic state such as Gardnerella and Prevotella also benefit from glycogen to grow to high numbers, produce biogenic amines (odor), cause inflammation (recruiting target cells for HIV to the mucosal surfaces), desialylation of the mucosa and exfoliation of the vaginal epithelium (discharge). Lower glycogen stores may partially explain why certain women with this microbial state have less symptoms, such as odor, discharge and inflammation, than other women.
Background During reproductive years glycogen is synthesized by the tissues of the reproductive tract [3, 4] and is essential for early embryo implantation and development. Glycogen shed into the vaginal lumen functions as a carbon source for vaginal lactobacilli such as Lactobacillus crispatus [5, 6] and iners but also for Gardnerella vaginalis and Prevotella bivia (unpublished data). Glycogen levels are reduced in women who have Lactobacillus-depleted microbial states [7] which could be explained by the interplay between host factors (glycogen synthesis) and bacterial factors (glycogen breakdown) [8].
It has been well-established that ethnicity is an important factor determining the odds of a woman to have bacterial vaginosis. Women of African descent are found to have more Lactobacillus iners than Lactobacillus crispatus and are more often colonized by a Lactobacillus-depleted community [9-11]. These women often have symptoms such as discharge and odor, and are at higher risk of acquiring sexually transmitted infection [12, 13], are more likely to have persistent HPV colonization [14, 15] and are more likely to give birth prematurely [15-17].
There has been considerable effort to understand the variation of health outcomes and symptoms amongst women with comparable vaginal microbiota. Most studies were directed at understanding this variation from a bacterial perspective, for instance by looking at genetic variation amongst bacterial isolates of Gardnerella vaginalis [18, 19] and Lactobacillus crispatus [6, 20]. We believe that the dataset we propose to analyze provides a unique opportunity to also take host factors into consideration.
Variations in glycogen metabolism due to host genetic factors may affect both hepatic glycogen accumulation as well as endometrial and vaginal glycogen accumulation. For a long time it was thought that progesterone was the main controlling hormone for glycogen synthesis in the endometrium [21, 22]. A recent study has now found that the progesterone/glycogen link is only indirect. Insulin is the hormone that inactivates glycogen synthase kinase 3β thereby activating glycogen synthase, similarly in liver tissue as well as in endometrial cells [23].
These commonalities lead to our hypothesis that there may be a genetic relationship between bacterial vaginosis and type 2 diabetes. In genome-wide association studies into genetic risk factors for type 2 diabetes several SNP’s have been identified. The most robust and consistent SNP’s are found in the TCF7L2 gene. Certain SNP’s in this transcription factor (elsewhere referred to as a TCF/LEF or TCF7) confer up to twofold increased risk of developing type 2 diabetes [24]. TCF7L2 knockout mice had reduced glycogen stores in the liver [25]. People carrying the risk allele synthesized less insulin in response to a glucose challenge test [26, 27]. This lower insulin production may have consequences for glycogen accumulation in the reproductive tract. Less glycogen synthesized means less carbohydrate for vaginal Lactobacillus species to acidify and Gardnerella to cause symptoms, odor and discharge. As of yet the ethnic differences in type 2 diabetes (where African ethnicity are at higher risk) were not able to be traced back to any SNP’s. The link between one specific TCF7L2 SNP’s (rs7903146) and type 2 diabetes was found in various regions of the world.
This would be the first study into host genetic factors and vaginal colonization.. The strength that this analysis offers is that it does not only include people from European heritage but includes a variety of groups with Asian, African and European roots. Clues from this study may inform bigger studies where we could look at the influence of genetic factors on lactate and glycogen taking into account the different ethnic groups and vaginal colonization patterns. If glycogen metabolic commonalities are found between diabetes and BV, this opens up a wide range of new treatment options for BV and related conditions, and possibly novel applications for established diabetes drugs including amylase inhibitors, insulin and metformin.
Proposal
To study the SNP’s of the TCF7L2 gene and their relationship to vaginal microbiome. The HELIUS cohort will this year have both datasets available for 346 women. The vaginal microbiome of these women was previopusly studied and the authors are willing to share these data too. We propose to look at three SNP’s of the TCF7L2 gene, specifically rs7903146, rs 4506565 and rs12255372 and study whether their relationship with having a Lactobacillus-deplete (case) or Lactobacillus-dominated (control) microbiome. In case the study cohorts permit (and the odds ratio), we could differentiate further into ethnicity. In case we find a strong signal that would warrant further study, we could look at glycogen concentration, lactate concentration, vaginal microbiome and ethnicity. There are many more vaginal swabs available and SNP data for 12,000 participants in total.
Collaboration and transparency This is an Open Kitchen Science project, meaning that the aim is to maximize collaboration and transparency. All data files, methods and results, either positive or negative, will be released and published prior to peer review (either on a preprint-server or on blog). Regular information about proceedings will be shared (including this proposal, without financial details) on blogs.
1. Snijder, M.B., et al., Cohort profile: the Healthy Life in an Urban Setting (HELIUS) study in Amsterdam, The Netherlands. BMJ Open, 2017. 7(12): p. e017873.
2. Borgdorff, H., et al., The association between ethnicity and vaginal microbiota composition in Amsterdam, the Netherlands. PLOS ONE, 2017. 12(7): p. e0181135.
3. Milwidsky, A., Z. Palti, and A. Gutman, Glycogen metabolism of the human endometrium. J Clin Endocrinol Metab, 1980. 51(4): p. 765-70.
4. Mirmonsef, P., et al., Glycogen Levels in Undiluted Genital Fluid and Their Relationship to Vaginal pH, Estrogen, and Progesterone. PLoS One, 2016. 11(4): p. e0153553.
5. Hertzberger, R.Y., A. Brandt, and R. Kort, Carbohydrate active enzymes in Lactobacillus crispatus – a possible link between the pullulanase gene and growth on glycogen. Figshare, 2018.
6. van der Veer, C., et al., Comparative genomics of human Lactobacillus crispatus isolates reveals genes for glycosylation and glycogen degradation: Implications for in vivo dominance of the vaginal microbiota. bioRxiv, 2018: p. 441972.
7. Mirmonsef, P., et al., Free glycogen in vaginal fluids is associated with Lactobacillus colonization and low vaginal pH. PloS one, 2014. 9(7): p. e102467.
8. Vaneechoutte, M., The human vaginal microbial community. Res Microbiol, 2017. 168(9-10): p. 811-825.
9. Fettweis, J.M., et al., Differences in vaginal microbiome in African American women versus women of European ancestry. Microbiology, 2014. 160(Pt 10): p. 2272-2282.
10. Ma, B., L.J. Forney, and J. Ravel, Vaginal microbiome: rethinking health and disease. Annual Review of Microbiology, 2012. 66: p. 371-389.
11. Ravel, J., et al., Vaginal microbiome of reproductive-age women. Proceedings of the National Academy of Sciences of the United States of America, 2011. 108 Suppl 1: p. 4680-4687.
12. Gosmann, C., et al., Lactobacillus-Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity, 2017. 46(1): p. 29-37.
13. Sewankambo, N., et al., HIV-1 infection associated with abnormal vaginal flora morphology and bacterial vaginosis. Lancet (London, England), 1997. 350(9077): p. 546-550.
14. Kero, K., et al., Association of asymptomatic bacterial vaginosis with persistence of female genital human papillomavirus infection. Eur J Clin Microbiol Infect Dis, 2017. 36(11): p. 2215-2219.
15. Brown, R.G., et al., Establishment of vaginal microbiota composition in early pregnancy and its association with subsequent preterm prelabour rupture of the fetal membranes. Translational Research, 2018.
16. Donati, L., et al., Vaginal microbial flora and outcome of pregnancy. Archives of Gynecology and Obstetrics, 2010. 281(4): p. 589-600.
17. Martius, J. and D.A. Eschenbach, The role of bacterial vaginosis as a cause of amniotic fluid infection, chorioamnionitis and prematurity–a review. Archives of Gynecology and Obstetrics, 1990. 247(1): p. 1-13.
18. Schellenberg, J.J., M.H. Patterson, and J.E. Hill, Gardnerella vaginalis diversity and ecology in relation to vaginal symptoms. Res Microbiol, 2017. 168(9-10): p. 837-844.
19. Janulaitiene, M., et al., Phenotypic characterization of Gardnerella vaginalis subgroups suggests differences in their virulence potential. PLoS One, 2018. 13(7): p. e0200625.
20. France, M.T., H. Mendes-Soares, and L.J. Forney, Genomic Comparisons of Lactobacillus crispatus and Lactobacillus iners Reveal Potential Ecological Drivers of Community Composition in the Vagina. Appl Environ Microbiol, 2016. 82(24): p. 7063-7073.
21. Jaffe, R.C., D.M. Stevens, and H.G. Verhage, The effects of estrogen and progesterone on glycogen and the enzymes involved in its metabolism in the cat uterus. Steroids, 1985. 45(5): p. 453-62.
22. Mimori, H., et al., Effect of progestogen on glycogen metabolism in the endometrium of infertile patients during the menstrual cycle. Fertil Steril, 1981. 35(3): p. 289-95.
23. Flannery, C.A., et al., Insulin Regulates Glycogen Synthesis in Human Endometrial Glands Through Increased GYS2. J Clin Endocrinol Metab, 2018. 103(8): p. 2843-2850.
24. Florez, J.C., et al., TCF7L2 polymorphisms and progression to diabetes in the Diabetes Prevention Program. N Engl J Med, 2006. 355(3): p. 241-50.
25. Boj, S.F., et al., Diabetes risk gene and Wnt effector Tcf7l2/TCF4 controls hepatic response to perinatal and adult metabolic demand. Cell, 2012. 151(7): p. 1595-607.
26. Jainandunsing, S., et al., Transcription factor 7-like 2 gene links increased in vivo insulin synthesis to type 2 diabetes. EBioMedicine, 2018. 30: p. 295-302.
27. Loos, R.J., et al., TCF7L2 polymorphisms modulate proinsulin levels and beta-cell function in a British Europid population. Diabetes, 2007. 56(7): p. 1943-7.
Power to “the crispies” – what I learned at the Keystone symposium on the genital microbiome.
In December I visited the long awaited Keystone symposium on the Role of the Genital Microbiome in Reproductive and Sexual Health. It was such an exciting and interesting conference and I learned a lot. Even in the airplane I spent the very last minutes of the flight talking about vaginal bacteria.
The strength of this symposium was the diversity. There were doctors and lab people, students and professors, there were Africans, Europeans, Americans and Asians. But most importantly, they came from different fields: HIV, preterm birth, etc. These people normally would not be at the same conference, they live parallel scientific lives publishing in their own journals and attending their own meetings. But the bacteria of the vagina bring them together. It has become crystal clear that the vaginal flora (or “microbiota”) have a very strong say in the various questions of these fields, such as who is prone to HIV infection (sexual health) and who is at high risk to deliver their baby too soon (reproductive health).
These people sat down in a big conference room together for four days, with in total 22 hours of presentations and about ten hours of more random wine-infused scientific chatter around 70 square feet of results printed on big poster boards.
The collective results were striking to say the least. One study that turned up in several presentations a group of 236 young South-African women were followed for about a year. Just as a reminder: in certain areas of Africa women start out with 0% HIV at 14 years old and in the following six years about 60% gets infected with HIV. In certain communities being seropositive is the norm. In this particular study, on the FRESH cohort in a township in Kwazulu Natal, a group of women were followed for about two years, 13% of all women got infected with HIV during this time period. But the risks of getting HIV-infected were not the same for all of them. The subgroup that had a vaginal flora mostly made up of a bacterium called Lactobacillus crispatus exactly 0 girls were infected. These bacteria seemed to almost behave like condoms (although please please still use condoms).
Find the paper here: https://linkinghub.elsevier.com/retrieve/pii/S1074-7613(16)30519-2
This same picture returned in literally every presentation that followed. Women with Lactobacillus species always have better health outcomes, and in most studies women with Lactobacillus crispatus do a bit better than Lactobacillus iners. The only exception I can think off is Candida infection that occurs regardless of the vaginal microbiome or in some studies a bit more often in women with vaginal lactobacilli.
Jeanne Marazzo, the opening speaker of the symposium, showed a picture of the vaginal lactobacilli a protective superpower. A dangerous watchdog that keeps out the bad guys. It was fascinating for me, yet again, to see this one Lactobacillus species beyond any doubt, in virtually every talk at the conference, in any cohort, be the good guy. I confirms that it is important and worthwhile to focus on these bacterial species because they mean so much for women’s health.
What happens if you don’t have these crispies (as I heard them refer to during the conference) or you lose them in the course of your life? Well, one of the most prevalent vaginal bacteria is a species called Lactobacillus iners (either pronounced like “ainers” pronounced like “I nurse” or “inners” like in “inner circle”). Clearly, this is the less preferred Lactobacillus. A bacterium with two faces: women that have this bacterium as their most abundant species seem less well protected and more likely to get infected or when pregnant deliver their babies too early. However, Lactobacillus iners is still preferred over having no Lactobacillus at all. You could see iners as just not a very good watchdog. Maybe he doesn’t bite? Or maybe he is secretly friendly with the bad guys? Or maybe he is just careless: leaving windows and doors open to let the bad guys in? All questions up for debate.
Douglas Kwon, who was the senior author on the previously mentioned study on HIV, showed data that sketch a rough storyline of vaginal microbiome. Most women start out with “crispies”, if they change to a different bacteria, it’s most often iners, and if they change from iners their more likely to switch to a flora that has very few lactobacilli but lots of other unwanted species such as Gardnerella, Prevotella and Megaspaera. These bacteria are not regarded as real bad guys (like Salmonella or Chlamydia) that will directly make you sick, although some believe they are. Clearly, you still really need to have unprotected sex with an infected partner before you get infected with Chlamydia or HIV.
Having no lactobacilli but these bacteria like Gardnerella is regarded as a “state” and referred to as “dysbiosis”. Although in many African groups it is more common than having Lactobacillus. Most women wouldn’t even notice that they don’t have good watchdogs around but some have symptoms like discharge and odor. For some it is so serious that it just outright destroys their sex lives, their relationships and their confidence. Even without preterm birth and HIV, this would already be a pretty good reason to study it and try to help these women out.
The immune response
One of the things that I learned more about during the conference is the role of the immune system. Without your lactobacilli, the immune system is more likely to be alarmed. It starts to get ready to attack and reacts as if its fighting off an infectious disease (we call this inflammation: redness, swelling, pain caused by your immune system responding to an infection). This is bad news if you have sex with a partner infected with HIV because HIV viruses infect exactly those cells (T-cells) that are involved in the immune response. So basically, you are calling the victims to the party.
When we talk about the immune system we distinguish between “pro-inflammatory” and “anti-inflammatory” signals: bacteria or chemicals that are the equivalent of someone yelling “fire fire!” to the immune system (pro-inflammatory), or sing a lullaby to the immune system (anti-inflammatory). Both may be going in in the vagina. Species like Gardnerella and Prevotella may already on their own may send alarming signals. And chemicals produced by Lactobacillus may be the equivalent of the lullaby: a comforting signal to the human body that “the good guys” are around.
When your pregnant the story may be a bit different. It could be that these bacteria alone may cause your water to break way too early and give your baby a troublesome start (or worse). One of the mechanisms through which this happens may be inflammation. Several stories in the symposium outline how Lactobacillus species will prevent an inflammation reaction whereas species like Gardnerella will provoke inflammation. Some of the pathways through which our bodies start such an inflammatory response are the same that the pregnant body uses to start labor.
Before I move on to my own research let’s just sit back and look at the weirdness of the vagina. How incredible it is that women really only just have two important watchdogs. In some groups you will find about 80% of women having one of these. This is found nowhere else in the human body and as Jeanne already pointed out: we really need to get rid of “diversity” as a characteristic of a healthy flora (or “microbiome”). In the vagina you want less species not more.
It was really great to meet so many of the heroes of the field. Everyone took a lot of time to answer all questions and everyone was very approachable.
Unfortunately, I cannot share many of the very interesting new aspects of the vaginal microbiome that were presented during the meeting, because a lot of the results are unpublished.
If you want to see the poster I presented, please find it here: https://figshare.com/articles/Lactobacillus_crispatus_growth_on_glycogen_is_dependent_on_its_type_1_pullulanase_gene_variant/7478330
I also gave a 10 minute oral presentation and I used my slides later for a lecture at ARTIS/Micropia. Find the presentation here: https://figshare.com/articles/Introduction_to_vaginal_microbiome_-_lecture/7583312
What’s next?
This year I am planning to make much more time for research. I want to further look at glycogen metabolism of vaginal bacteria and luckily I have met several people at the conference who are working on the same topic and are interested to work together.
The meeting really stresssed the importance of trespassing the boundaries of different fields. I really want to pull the human factor into the lab. Study the presence of glycogen levels and glycogen breakdown enzymes in vaginal swabs. Also, I will continue studying the type 1 pullulanase of L crispatus and regulation of this gene/enzyme. Having seen the talks in Cape Town I think that these basic mechanisms of vaginal metabolism need to be elucidated so we can move on. Deborah Jekel, a very motivated and talented bachelor student is starting this week to help me do just that.
Stay tuned!
News!
Great news! We sent an abstract to the Keystone conference “Role of the Genital Tract Microbiome in Sexual and Reproductive Health” and it was selected for a short talk. So I’ll be presenting the research on stage in Cape Town in December!
This is the abstract:
Lactobacillus crispatus growth on glycogen is dependent on its type 1 pullulanase gene variant.
Rosanne Y. Hertzberger1,2, Alicia Brandt 2,3, Charlotte vd Veer4, Jorne Swanenburg1,5, Remco Kort1,5
1 VU University Amsterdam, Netherlands, 2 REBLAB, Netherlands, 3 University of Groningen, Netherlands, 4 Public Health Service, Amsterdam, Netherlands, 5 TNO, Zeist, Netherlands
Glycogen is an abundant carbohydrate in the vagina of reproductive-age women. Levels of free glycogen vary depending on the makeup of the vaginal microbiota: women with bacterial vaginosis, a common dysbiosis with higher pH and few lactobacilli, generally have lower glycogen levels. This finding has led to the general assumption that lactobacilli depend on this glycogen and use it as a carbon source for colonization and acidification. However, direct evidence for this mechanism has been lacking sofar.
We studied growth, enzymatic activity and metabolite production of a group of 19 Lactobacillus crispatus strains that were isolated from reproductive age women. 13 out of these 19 isolates were able to grow on glycogen whereas 6 did not use glycogen for growth. Starch was used as a proxy to study glycogen breakdown and starch metabolism activity was found in both supernatant as well as in the washed pellets after growth on glycogen. Several strains did not show activity after growth on glucose indicating differential regulation of this metabolic pathway.
A survey of the genomes to track down any carbohydrate active enzymes showed the presence of a putative cell surface associated type 1 pullulanase. The gene is predicted to encode a 140 kDa enzyme consisting of an N-terminal signal peptide, two carbohydrate binding modules, a carbohydrate active module belonging to the GH13_13 family and a C-terminal S-layer associated protein (SLAP) domain. The gene shows high similarity to the extracellular cell wall attached pullulanase previously found in a human gut isolate of Lactobacillus acidophilus.
Although a copy of the pullulanase gene was found in the genomes of all 19 L. crispatus strains, the six non-glycogen consuming strains all carried one of three different mutations in the N-terminal signal peptide sequence, expected to disrupt transcription of at least part of the gene.
Our results show for the first time that certain vaginal isolates of Lactobacillus crispatus are capable of metabolizing one of the most abundant vaginal carbohydrates. We identify a pullulanase that may be essential for this activity. These findings bring us further to understanding the basic mechanisms of Lactobacillus colonization and acidification of the human vagina.
This project is part of REBLAB, an Open Kitchen Science initiative. For more information see www.reblab.org.
Also: a paper with the glycogen results (reproduced in the TNO lab) is under review at Microbiome journal and available in the meanwhile at bioRxiv, find it here https://www.biorxiv.org/content/early/2018/10/13/441972
Observations…
Hi everybody,
This summer I did some preliminary test with 4 strains (2 glycogen consumers, RL_010 and RL_011, and 2 non-consumers RL_009 and RL_026) to see whether maltose is a suitable substrate to be able to compare activity between glycogen consuming and glycogen non-consuming strains of L. crispatus. I also used this experiment to get some additional replicates for Figure 3 of the bigger June blog post (this one). Will be updating this soon. First need to repurchase a GraphPad PRISM serial code 🙁
Preliminary observations of the experiment:
-something went wrong with the dilution of the culture for strain 9 (5 times instead of 10 times diluted), so I won’t be able to use this data point.
-the 2 glycogen consuming strains show starch degrading activity in both pellets as in the supernatants, like I saw before. (could this be the S-layer effect?)
-strains grow well on maltose but the activity is still somewhat repressed (or not induced?) just like on glucose. Next step is to try this on maltotriose to see if this may be a substrate that all strains will grow on and pullulanase activity will also be present.
-suprisingly, one strain (RL_011) does seem to induce (or fails to repress) the pullulanase activity on glucose and on maltose. Is there another level of variation in this aspect?
Experiment wishlist
Back in the lab! Hope to spend a lot of time here in the coming weeks. My program is getting busier in October so I hope I can get a lot done before that. On my experimental wishlist:
-Study Gardnerella vaginalis and Lactobacillus iners growth on glycogen and starch breakdown. For this I need to be able to create more strict anaerobic conditions then the growth protocols I am using now since these species seem to be more sensitive to oxygen. Let’s see if we can arrange that in the lab.
-Further compare glycogen consumers and “non-consuming” strains of L. crispatus. To be able to make this comparison I will need to find growth conditions in which I get expression of the gene, but growth of both consumers and non-consumers. Alicia told me that maltose, maltotriose or maltopentaose may do the trick. The precise mechanisms of this induction of expression (?, or whatever it may be) are interesting and may have in vivo relevance, but have low priority. I just want to compare the strains, but no growth (of the non-consuming strains on glycogen) means no pellet and no bacteria to use as a comparison! I will start with maltose, and have ordered maltotriose. These experiments will also be useful to get more replicates for the growth data in the previous blog.
-Alicia also told me more about the differences between amylose and amylopectin in starch. This may explain why we only saw partial breakdown of starch in activity assay of L. crispatus. If we are able to differentiate between amylose and amylopectin utilization, we may also learn more about the exact activity and ways to use this as a target for therapeutic approaches.
-At some point I really need to make an overview on what is actually known about glycans in the human vagina. What techniques and stains were used in the different studies sofar to characterize the glycans?
Stay tuned!
Carbohydrate active enzymes in Lactobacillus crispatus – a possible link between the pullulanase gene and growth on glycogen
Hello everyone, after moving to Rotterdam and having a baby (welcome Frank!) I am back here to show you the progress I made! The findings I present here are work in progress, but it is time to give a first update. I believe the results I present here are promising on three different levels: 1) content-wise, I think they are a start to understanding glycogen metabolism of Lactobacillus crispatus better 2) I am showing it all: data, methods, results, questions and flaws. Exactly how I envision Open Kitchen Science, (although with a bit of delay, so less “realtime” as I would like) 3) this is the work of a collaboration with another “postacademic” scientist, who wants to contibute to science besides her regular job. Meet Alicia Brandt!
Before diving into the science, a few points of order:
Collaboration: As mentioned before, I am very interested in collaborating and receiving feedback on my blog posts. Please contact me, or leave a remark. I am also interested whether any other labs are working on the same question or have been working on this in the past. If you are (or were ) doing experiments with Lactobacillus crispatus we could find out whether these results are true for more strains (especially using the same medium). Let’s talk! Twitter or Facebook or email rosanne dot hertzberger at gmail dot com
Manuscript: Part of the results presented here will be part of a paper that is currently in preparation by Charlotte van der Veer and Remco Kort and co-authors (including me). I will update this blog post as soon as this paper is published. When the paper is published, the full genomes of the strains will also be released online.
Internship: starting after the summer break I hope to be supervising a student working on this project, together with Jurgen Haanstra at the VU University Amsterdam. Find the description here (lab-site) or here (FigShare). BSc or MSc, please contact me if you are interested!
Warning: The results presented in this blog post are unreviewed and have not been replicated by an independent laboratory.
Open data All data and protocols used can be found in this FigShare collection.
How to cite: Hertzberger, Rosanne; Brandt, Alicia (2018): REBLAB: Carbohydrate active enzymes in Lactobacillus crispatus – a possible link between the pullulanase gene and growth on glycogen. figshare. Collection.
https://doi.org/10.6084/m9.figshare.c.4133819.v3
Short overview of where we are.
I am interested in glycogen metabolism of vaginal microbes. Glycogen is an abundant vaginal source of carbohydrates and varies depending on the vaginal bacterial signature. Roughly spoken: when the vaginal microbiota are dominated by Lactobacillus glycogen is generally higher. One of the overarching questions in the field is whether vaginal lactobacilli can metabolize this glycogen to grow and produce lactic acid. I think this is a pretty big deal. I have had a Google Scholar Alert with the keywords “vaginal glycogen” since a few years and not a week goes by without a paper getting published talking about how this vaginal glycogen is supposedly turned into lactic acid by lactobacilli. In my view, there is no evidence to back up this statement.
During my postdoc at Washington University St. Louis it became clear that the BV-associated Gardnerella vaginalis and Prevotella bivia are capable of growing on glycogen, as well as Lactobacillus iners (often encountered in vaginal microbiota with and without BV). Previously on this blog I showed experiments that Lactobacillus crispatus can grow and acidify using glycogen as a source. Here we zoom in further on the variation in glycogen metabolic capacity of different Lactobacillus crispatus strains, using a group of 23 different Lactobacillus crispatus isolates.
Summary of the findings
A dbCAN survey of the genomes of these strains shows an overview of several carbohydrate-active enzymes possibly involved in glycogen breakdown. One of the genes present in all strains is a putative S-layer linked type 1 pullulanase (GH13_13). Zooming in on this gene it appears that there is strong variation in the N-terminal sequence encoding a putative signal peptide. Only the strains that do not have a mutation in the sequence of this signal peptide show growth on glycogen. 6 out 23 strains that have a mutation in this peptide cannot grow on glycogen. Is this signal necessary for Lactobacillus crispatus to break down the big molecules of glycogen in the vaginal environment? Nothing final yet, but a promising lead to look further into.
Alicia Brandt: dbCan analysis and glycogen-active enzymes
Since I started with REBLAB I encountered several people who shared their own stories about their departure from science and how this affected them. Sometimes these people find the time and energy to still contribute to science parallel to their regular jobs. We start to call ourselves “post-academic” scientists. One of these people is dr. Alicia Brandt (previously Alicia Lammerts van Bueren as she is known in the glycobiology world) who left science last year for a job in a supporting role at the Young Academy at Groningen University. She expressed the strong wish to keep being involved and to share her knowledge and skill. It’s a happy coincidence that her expertise is exactly what I am looking for: glycobiology and even glycogen metabolism of bacteria. I went to meet her in Groningen and had a great time!

Alicia and me on the steps of the Groningen academy building
Lactobacillus crispatus isolates in this blog
Last year I was very lucky to join Prof. dr. Remco Kort at the VU University Amsterdam who had just isolated and sequenced about 30 Lactobacillus crispatus strains from women with and without BV ecology. These women were patients at the GGD facility in Amsterdam and the strains were isolated as part of the thesis work of Joke Dols. MSc student Jorne Swanenburg was responsible for the genomic analysis sofar and Charlotte van de Veer is currently finalizing her thesis and is writing up a paper on carbohydrate metabolism of these strains. Recently, a paper was published where the strains were used as a reference for a possible new vaginal prebiotic. The paper is open access and the isolation is described in materials&methods. At the moment we are analyzing these strains, some appear harder to culture than others. The list of isolates that we are able to maintain in the lab is a work in progress and will be updated later. We were unable to revive RL_005 from its glycerol stock and the sequencing file of RL_022 has some problems.
Alicia offered to use her expertise to help answer some questions surrounding Lactobacillus crispatus glycogen metabolism. First thing she did was to perform an analysis using the so-called dbCAN server, detecting the presence of several glycoproteins in the genomes of a set of isolated Lactobacillus crispatus strains that can potentially be involved in glycogen metabolism.
Find the raw data here (FigShare).
Find the protocol here (FigShare).
Most important findings:
- All L. crispatus genomes contained a putative cell-surface associated (as indicated by SLAP domain) pullulanase Type 1 enzyme (http://www.uniprot.org/uniprot/A0A135Z466) implicated in glycogen degradation. Key features of this enzyme are the presence of a CBM41 and CBM48 and a GH13_13 catalytic domain (Fig 1). More on this gene/protein later in this blog post.
- There is a cluster (operon?) of alpha-glucan degrading enzymes in all crispatus genomes which include a GH65 (phosphorylase?), GH13_20(CBM34) and GH13_31-2 enzyme. Further analysis required to see if they are co-transcribed. (Table 1, genes located in the cluster are indicated with an (a)).
- Other alpha-glucan metabolizing enzymes found within the L. crispatus genomes include: GH13_18, GH13_29, GH13_31, GH31 (see Table 1).
- Only three strains of L. crispatus contained a GH13_18 (RL02, RL09, RL10) (Note: RL_022 contained this enzyme as well, but sequencing file of strain has errors).
- RL_006 is the only strain that does not contain a GH13_29 enzyme.
Table 1: Overview of Alpha-glucan enzymes found in L. crispatus genomes (see www.cazy.org for more info on predicting enzyme activities based on amino acid sequence similarities with known enzymes within a given family)
| Enzyme Family | Proposed Activity | Genomes |
| GH13_13 (CBM41, CBM48) | Pullulanase type I | All except RL31, RL32 |
| GH13_18 | Sucrose phosphorylase | Only RL02, RL09, RL10. RL22* |
| GH13_20 (CBM34)(a) | Pullulanase type III, cyclodextrinase | All |
| GH13_29 | Trehalose-6-phosphate hydrolase | All except RL06 |
| GH13_31-1 | oligo-alpha-1,6-glucosidase | All |
| GH13_31-2 (a) | oligo-alpha-1,6-glucosidase | All |
| GH31-1 | alpha-glucosidase | All |
| GH31-2 | alpha-glucosidase | All |
| GH65 (a) | Maltose phosphorylase | All |
*RL22 genome needs to be resequenced (problems with sequencing file)
(a) Constitutes part of a cluster of enzymes, possibly an operon.
N-terminal signal peptide of the type 1 pullulanase gene (GH13_13) corresponds with growth on glycogen.
Glycogen is a prevalent potential carbon source in the vagina of reproductive age women. Lactobacillus crispatus and Lactobacillus iners-are the most frequently encountered species vaginally. They are generally assumed to be responsible for the low pH and high lactate concentrations- but it is unclear what sugar source they use for lactic acid production.
(To be fair: there are many assumptions and uncertainties here. To name a few: there is only circumstantial evidence that vaginal lactic acid is of bacterial origin. The same counts for the human origin of vaginal glycogen. Actually, the fact that we are dealing with glycogen and not with a different glycan is not all that well established since most studies used a PAS stain and an alpha-glucosidase. It could well be that we are dealing with a different glycan. However, I am cutting a few corners here and will assume that lactate is of bacterial origin and the glycan is in fact glycogen, produced by the hostess herself.)
Previously it was reported that at least a subset of vaginal lactobacilli (jensenii, gasseri and johnsonii) were unable to breakdown glycogen, but this study did not look at glycogen metabolism of crispatus and iners. I presented on this blog some evidence that the DSM strain of L crispatus is capable of growth and acidification (producing lactic acid) on glycogen as the carbon source.
This possible L. crispatus glycogen metabolism could be an important player in the acidification of the human vagina, and the health benefits that are associated with a vaginal community dominated by L. crispatus. Needless to say, we would like to know more about it.
Why the type 1 pullulanase had my interest – the role of serendipity
This gene had had my attention already since I started working with a set of 4 Lactobacillus crispatus strains during my postdoc in St Louis at the Lewis lab, WUSTL School of Medicine. These strains were MV-1A-US, MV-3A-US, JV-V01 and 125-2-CHN. I found that two of these (MV-1A-US and MV-3A-US) were able to grow on glycogen as a carbon source whereas two others (JV-V01 and 125-2-CHN) were not. To find a possible genetic origin of these differences I did a blastp analysis using several enzymes as a query that were known to be involved in glycogen (or starch) breakdown. Those were: glgX of E. coli, sap of Streptococcus agalactiae (see paper and sequence ), SusB of Bacteroides theta and the amylase (amyE) of Bacillus subtilis. Three out of the four genes showed no full-length copies in the four L. crispatus genomes, but I did find genes similar to the glgX gene: a gene annotated as a type 1 pullulanase (uniprot link of the copy in strain MV-3A-US). One of the strains unable to grow on glycogen (125-2-CHN) had no copy of this gene and the other strain unable to grow on glycogen (JV-V01) had a mutation in the upstream region, that I thought might be a dealbreaker for expression. This paper on comparative genomics of Lactobacillus crispatus confirms the absence/presence of the type 1 pullulanase in these strains. (supplementary material file nr 5). All other strains in the comparison contained the gene except for 125-2-CHN and 214-1.
This was all purely speculative at that moment, but this is why the type 1 pullulanase gene caught my attention: it was the only one that showed clear variation amongst the L. crispatus strains. There was a big chunk of serendipity that lead me to the presented finding.
All L. crispatus strains have the type 1 pullulanase gene, but the N-terminus looks different!
I was therefore disappointed that initially, from Alicia’s dbCAN analysis, and also in a screen performed by Jorne Swanenburg, it became clear that all strains had a copy of this gene. I further analysed this gene and also included the upstream region (expecting to find a mutation similar to the one in the JV-V01 strain). The sequence directly upstream this gene encodes a putative signal peptide (see Figure 1) and there is strong variation amongst the collection of genomes in this particular area (see Figure 2)
Please find the genes from all strains here. I used the EMBL Clustal Omega online Multiple Sequence Alignment tool to compare the genes and the results were striking. You could redo the analysis by using the file and just copy paste it into the Clustal Omega tool. The genes are conserved but not the starting region. In Figure 1, a schematic overview is shown of the organization of this gene in L. crispatus. Thanks Alicia!) In Figure 2, I am showing the variation in the N-terminal sequence from the aforementioned Clustal Omega comparison.
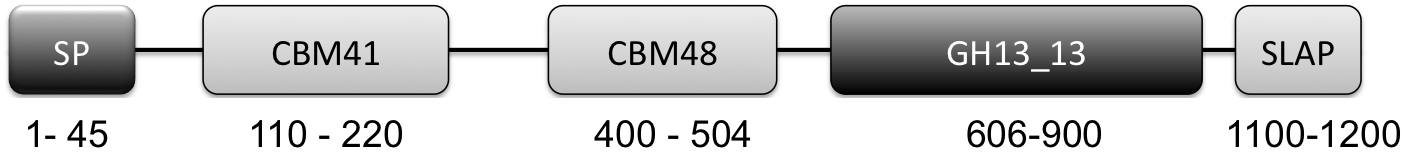
Figure 1: Graphical Representation of GH13_13 Pullulanase Type 1. (N to C terminal): SP: signal peptide (amino acids 1-45), CBM41: carbohydrate-binding module family 41 (amino acids 110-220), CBM48: carbohydrate-binding module family 48 (amino acids 400-505), GH13_13: glycoside hydrolase family 13 subfamily 13 (amino acids 606-900), SLAP: Surface layer associated domain (1100-1259)
 Figure 2: Comparison of the N-terminal sequence of the pullulanase gene in L. crispatus strains. Adjusted from Clustal Omega. Red: strains with a possible disrupted N-terminal sequence and signal peptide. Blue: strains with an N-terminal sequence indicating an intact signal peptide. Find full sequences here.
Figure 2: Comparison of the N-terminal sequence of the pullulanase gene in L. crispatus strains. Adjusted from Clustal Omega. Red: strains with a possible disrupted N-terminal sequence and signal peptide. Blue: strains with an N-terminal sequence indicating an intact signal peptide. Find full sequences here.
7 out of the 24 strains (RL_#-strains) have a mutation in the N-terminal locus of the pullulanase gene, more specifically, in the sequence of the signal peptide. Those strains are indicated by red in Figure 2. Further experiments should indicate whether this means that the start site of these “red” strains lies more downstream from the start site in the “blue” strains.
Remarkably, there are five different variants of this gene locus present in those seven strains. For instance, strains RL_002 and RL_009 only show a deletion of two nucleotides (a frame shift), whereas strain RL_006 and RL_007 have a completely different sequence in this region.
Growth on glycogen of the L. crispatus strains.
I performed a very straightforward cultivation experiment using glycogen as a carbon source. Initially, I only used four isolates (RL_002, RL_003, RL_007 and RL_026). When these results were promising I started to screen all 23 strains . As a benchmark I used the DSM strain, which I previously showed is able to breakdown glycogen for growth and lactic acid production. I used the same methods as described in that blog post:
I inoculated the strains in regular NYCIII glucose and after ~72 hours of growth diluted them with NYCIII glycogen (final concentration 0,5%), water (as a negative control) and glucose dissolved in water (final concentration 0,5% as a positive control). After 48 hours I measured the optical density at 600 nm to determine growth on glycogen compared to growth on NYCIII without supplemented energy source and NYCIII glucose.
For 19 out of the 23 strains tested (we could not revive strain RL_005) I have either biological triplicates or duplicates, I am showing their results in Figure 3. I am still working on getting all data complete and aim to have at least two replicates of this experiment for each strain. In one biological replicate of strain RL_019 and one replicate of RL_006 results are very different from the others. I have no idea why and no ‘reason’ to exclude it. Not really sure what to do with this measurement at this point. All individual biological replicates are shown in the figure.
Note: I measured the cell density 10x diluted in PBS as well as undiluted. I am showing the diluted data in this blog post, since I do not have the undiluted culture measured on all dates for all strains. The difference between “growth” and “no growth” is more pronounced in the undiluted measurement since background absorption of the media is lower. I include an overview of the undiluted data in a separate tab of the excel sheet and the figure in the GraphPad Prism file. Other data that are in the file but not in the Figure shown here: the optical density in the positive (glucose) and negative control (water). Find the data here (.xls and the GraphPad Prism 7 file to generate the figure).
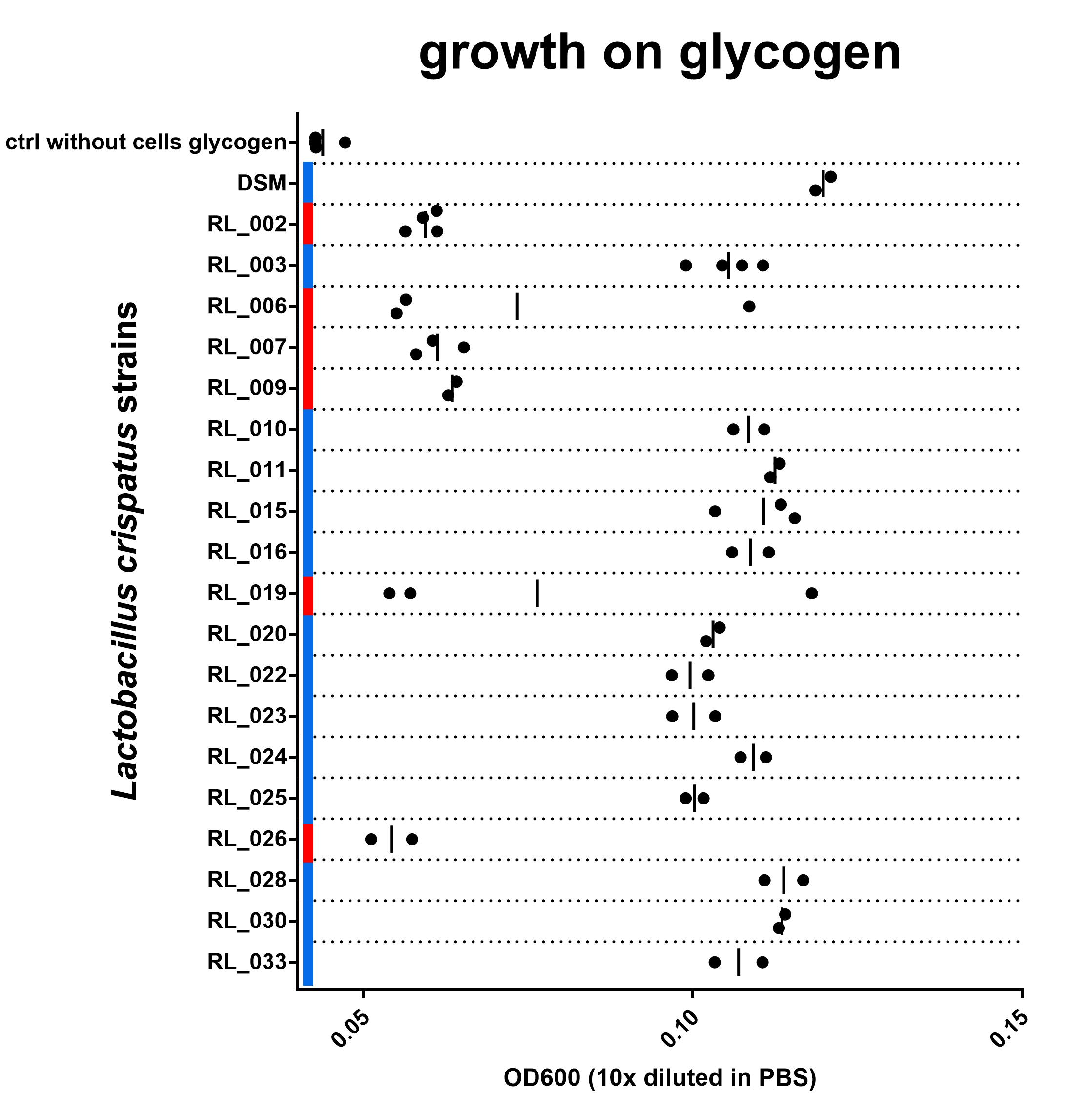
Figure 3: OD600 after culturing on NYCIIImedia with 0,5% glycogen. Biological replicates are shown as individual data points (some duplicates, some triplicates), vertical line indicates mean. Red or blue corresponds with red and blue in Figure 2. blue = with intact N-terminal signal peptide, red = with a disruption in this sequence. Find data here and protocol here
Although the growth data set is not complete yet, I do think we are seeing a strong connection between the N-terminal signal peptide of the type 1 pullulanase and the ability of the L. crispatus strain to grow on glycogen. All 6 strains in this experiment that have a disruption in the N-terminal signal peptide of the pullulanase gene show no growth on glycogen. The 14 strains that do have an intact signal peptide in the pullulanase gene can use glycogen as a source for growth. I see these data as strong evidence for an essential role of the pullulanase gene for glycogen consumption in Lactobacillus crispatus and, more specifically, the N-terminal signal peptide.
Thoughts, questions and new experimental plans
This finding is just that, a finding. Nothing final yet, an experiment that leads to a hypothesis: the N-terminal 29 amino acids are somehow important for Lactobacillus crispatus glycogen consumption. But how? Does this signal peptide lead to secretion of this enzyme? And if so, what is the influence of the C-terminal SLAP-domain? Are both required to localize this enzyme on the outer cell wall to be able to break down the big molecules of glycogen in its surroundings? Or does this signal peptide have a different function?
It is possible that the pullulanase without the signal peptide still functions in an intracellular metabolic pathway for glycogen breakdown, whereas the pullulanase gene that is localized on the outer cell wall can also debranch external glycans and utilize them for growth and lactic acid production.
I envision a few experiments to test these questions:
- it is probably important at this point to establish that the pullulanase is indeed a pullulanase. I have talked with Alicia about expressing the gene in E. coli to further characterize its activity. Alternatively, we could try to capture its native activity using cell free extracts and analyse carbohydrate products with Thin Layer Chromatography. Previously, we were able to show starch breakdown, next we should take a better look at the actual breakdown products.
- How can we study the role of the signal peptide? This is not that straightforward. Optimally we would just make L. crispatus mutants with and without it and track the enzyme’s activity and location. However, I have not seen any Lactobacillus crispatus cloning anywhere and not looking forward to try to develop my own protocols to get these isolates genetically accessible. (if someone has an idea, let me know). I hope to come up this summer with some experimental plan to test the cellular localization of this enzyme with and without the signal peptide.
What is the role of Lactobacillus crispatus glycogen metabolism in the context of the vaginal environment?
I believe that these experiments show that this activity is not something essential: the non-glycogen consumers seem to live happily in the vagina. Other lactobacilli, such as jensenii and gasseri also flourish in the vagina without being able to break down glycogen. Either, these bacteria utilize an alternative energy and carbon source. Or -what I personally expect- is that these bacteria live alongside glycogen consumers such as Lactobacillus iners or crispatus strains. Currently we don’t know whether the lactobacilli we encounter so abundantly in the vagina of reproductive age women are a collection of various strains or are a clonal population. It could well be that the non-glycogen consumers only thrive as freeloaders and can only colonize alongside a second glycogen-consuming species that does some of the glycogen breakdown. These exciting questions are definitely on my experimental wish list.
As I am still at home with baby, I will mostly do some reading and computer work to understand the genetic context and role of this pullulanase in other species. I found some interesting literature on this enzyme in Lactobacillus acidophilus and in a thermophilic bacterium called “Caldicellulosiruptor kronotskyensisencodes” (WOW! That must be one really interesting species!) Generally, pullulanases (and definitely secreted pullulanases) are of industrial relevance so there should be quite some protocols and knowledge out there. Also, I really need to read up on S-layers in lactic acid bacteria. Stay tuned!
And again: if you have suggestions how to continue, remarks or criticism, please let me know below. In general, I really appreciate any signs that this work matters to anyone because of the alternative publication route I am taking here.
Ruwe versie van tekst lezing Open Kitchen Science bij Spaarne Gasthuis “Wetenschap in Beeld” bijeenkomst.
Veel praten over Open Kitchen Science, weinig doen.Dat is een beetje de teneur van deze weken. Gelukkig ben ik ook bezig met een update over wetenschap. Ik ben over drie weken uitgerekend en hoop het voor die tijd af te hebben. Stay tuned! Hier een ruwe versie van lezing die ik een aantal keer heb gegeven de afgelopen tijd. Vind de bijbehorende slides hier.
Dames en heren, groot genoegen om hier voor u te spreken. Ik wil allereerst de organisatie bedanken voor de uitnodiging. Open Wetenschap en de manier waarop we wetenschap beoefenen is een belangrijk onderwerp en ik geloof dat er veel aandacht nodig is om de enorme verandering te bereiken die de wetenschap nodig heeft. Vandaag wil ik jullie mijn persoonlijke verhaal vertellen, mijn plannen en doelen met open wetenschap en het effect dat ik hoop dat het heeft op op reproduceerbaarheid, geloofwaardigheid, toegankelijkheid, kwaliteit en nu van wetenschap.
Een kort overzicht van mijn loopbaan tot dusver… (zie slide)
Helaas eindigde de tijd in een nogal zuur conflict met mijn begeleiders daar. Lang verhaal, lelijk voor hen, lelijk voor mij, lelijk voor wetenschap, maar het betekende dat ook al hadden wij een prachtig stuk wetenschap in onze handen, we toch zonder dat gepubliceerd te hebben naar huis ging.
Beide onderdelen van mijn carrière zijn erg belangrijk voor me, ik heb nooit willen kiezen, en mijn doel is ook om nooit te kiezen. De eerste jaren van mijn promotie en postdoc was het alleen zo dat wetenschap voor schrijven betaalde en nu is het andersom. Ik verdien geld met lezingen, schrijven, dagvoorzxitterschappen etc, en daarmee betaal ik voor de minder rendabele activiteiten: boeken schrijven, wetenschap.
Ik heb nooit willen kiezen en toch waren er momenten waarop de twee in conflict komen met elkaar en waarop ik toch de één moet voor laten gaan voor de ander. Dat gebeurde begin januari. Ik moest besluiten of het manuscript voor mijn boek op tijd af zou zijn om het voor de zomer te laten verschijnen, of dat ik alles uit mijn handen liet vallen en een aanvraag voor de Veni-beurs zou schrijven om te pogen alsnog een standaard academisch traject in te gaan.
Ik besloot voor mijn boek en schrijf carrière te kiezen. Tegelijkertijd vervulde het idee dat ik de wetenschap ging verlaten met een groot verdriet en gevoel van verlies. En ik ben opgegroeid met twee ouders, allebei arts, die mij leerden dat als een vraag je met een onmogelijk dilemma opzadelt, dat je alles in zijn werk mag stellen om antwoorden te vermijden, of in ieder geval zo lang mogelijk uit te stellen. Door de vraag ter discussie te stellen, of de context, of de onderliggende aannames. Dus dat deed ik. Het doel is om nooit te hoeven kiezen. Niet alleen life scientist, maar ook scientist for life. De vraag: wetenschap of geen wetenschap was té zwart-wit, en moest nodig ter discussie worden gesteld.
Dus besloot ik om met REBLAB te beginnen. Als “post academisch wetenschapper” met een radicaal transparante aanpak, oftewel Open Kitchen Science waarin het doel is om alles te publiceren dat los en vast zit: groot klein, positief negatief, leuk of niet leuk, high impact of low impact, methodes, posters, lezingen alles. Ik heb gastvrijheid aan de VU, in de groep van professor Remco Kort in het departement van Hans Westerhof, en heb dus toegang tot lab en VuNETID.
Beide aspecten van REBLAB, zowel Open Kitchen Science als post-academic wetenschapper zijn Nieuw terrain voor mij en vele anderen, en ik ben nog steeds bezig met alle aspecten ervan te ontdekken. Vandaag wil ik delen waarom ik deze route koos en de verwachte en onverwachte bevindingen.
Laten we het eerst over de voor- en nadelen van een standaard academisch traject hebben. De voordelen zijn duidelijk: beste baan in de wereld. De opwinding van het bestuderen van de chemie van het bestaan. Ik kan niets leukers bedenken en heb het altijd als een enorm privilege gezien.
Nadelen is de hele poeha er omheen. De cultuur, de dogma’s, het traditionele vastgeroeste rigide manier van communiceren. En misschien merk ik dat meer dan andere omdat ik elke week mag schrijven in NRC wat ik wil. Vrij, alleen, het is heerlijk om gewoon op send te drukken zonder eerst 7 co-auteurs te raadplegen en te wachten op de zegen van drie anonieme peers en dan na jaren je bevindingen in een onleesbare formele tekst in een obscuur blaadje te zien verschijnen. Waar je dan ook nog eens 3000 dollar voor moet betalen.
Maar dat publiceren is essentieel voor succes. In het academische monetaire systeem zijn high impact peer reviewed papers de munteenheid. En die papers moeten glanzen, vol staan met goed nieuws. Wetenschap ziet er altijd geweldig uit als het eenmaal is gepubliceerd. Chronologische logische volgorde. SLIDE
En omdat papers onze munteenheid zijn zie je veel geslotenheid. Huiver om te delen, je hoort mensen zeggen: wat als iemand ermee vandoor gaat? En dan vraag ik altijd: is dat dan niet de bedoeling van deze hele exercitie?
SLIDE Het grappige is dat je voor al mijn geschreven werk moet betalen. Het grote verschil is dat je bij NRC indirect mijn salaris betaaldt. En bij Elsevier Cell Host&Microbe moest ik juist zelf ook betalen om te publiceren. Dit is een enorm probleem in mijn ogen. Je kan dit zien als misbruik van publiek geld. Zowel de schrijvers als de lezers betalen en de reviewers ook, want die betalen met geld tijd en aandacht.
En het maakt me niet alleen boos als schrijver, als wetenschapper, maar ook als burger, als belastingbetaler. En als patient. Het publiek is hoger opgeleid dan ooit. Meer mensen gaan naar hoger onderwijs en komen in contact met wetenschappelijke ideeen, moeten papers lezen en soms ook schrijven. Vroeger dacht ik ook dat de mensen die toegang moeten hebben, die moeten weten hoe Lactobacillen H2O2 maken, dat die wel in een instituut zitten met toegang. Het is niet waar. Pas als je buiten de muren van de academie wordt gegooid zie je pas hoe hoog ze zijn. Hoe weinig zichtbaar je bent daar binnen.
Het maakt me verdrietig dat ik al meerdere keren papers moet opsturen naar de familie van patienten. Omdat jij wel toegang hebt en zij niet. Gelukkig is de open access trein al een tijdje op stoom. Mede dank zij initiatief van Nederland.
Maar ook met open access betaal je je nog steeds blauw aan publiceren. Gemiddeld artikel processing charges: 3000 dollar voor gouden standard toegang. SLIDE hier de prijzen van publiceren in de Plos tijdschriften. Daarnaast de andere publicatie-optie die ik nu onderzoek: een WordPress blog. Grote vraag is: in een tijd waarin iedereen alles gratis kan publiceren, wat is dan nog de toegevoegde waarde van uitgeverijen? Waarom zijn we bereid om ze zoveel geld te betalen terwijl er letterlijk honderden gratis publicaties opties zijn met gestoorde transparante peer review, die we ook wel een reactieforum noemen. Ik begrijp dat wat er in een officieel journal staat meer vertrouwen schept dan wat er op een obscuur forum plaatsen, maar is dat vertrouwen ook 3000 dollar per paper waard?
SLIDE Dit is wat universiteiten jaarlijks uitgeven aan publiceren.
SLIDE En deze communicatiedwangbuis is een belangrijke reden dat het er eigenlijk allemaal niet bijzonder aantrekkelijk uit ziet, dat standaard academische traject.
Het heeft bizarre consequenties. Bijvoorbeeld dat er nog steeds veel meer uitgeprint wordt dan we mogen zien op internet. Posters bijvoorbeeld. We laten elkaar best premature voorlopige resultaten zien van projecten, op papieren of textiel A0 formaat. Sommige mensen nemen stiekem foto’s ervan want je ziet die posters nooit online. Ze zijn alleen voor die paar duizend congresgangers, die er langs kunnen lopen. Waarom publiceren we ze niet? Waarom staan we er stiekem foto’s van te maken, of ze over te schrijven in een boekje? Het is 2018!
Nog een grappig effect: alles ter wereld kan je live volgen. Volvo ocean race racet door de zuidelijk ocean, kinderen van iedereen, oorlogsgebieden, de maan. Maar niet een praatje op een congres. Nee die kun je alleen volgen als er iemand in de zaal zit mee te typen. Live-tweeten van congres, hoe bestaat dat nog anno 2018?
Ik volg veel wetenschappers op Twitter. Ik weet hoe hun kinderen eruit zien, wat voor muziek ze van houden en waar ze op vakantie gaan. Maar zelden zeggen ze: hier is mijn wetenschap. Dit zijn de resultaten. Verse kennis, recht uit het lab. Het is echt ontzettend teleurstellend hoe weinig internet veranderd heft in de wetenschap. Sommige mensen zeggen dat er verandering op komst is, ik zie er heel erg weinig van. Dus toen ben ik het maar op eigen houtje anders gaan doen.
Daarom wil ik zo veel mogelijk delen. Vergelijk het met vloggers. Die mensen die urenlange beelden van de meest banale activiteiten. Er is een afkorting voor: TMI, too much information. Dat is precies hoe ik wetenschap wil doen. Ik wil elke methode publiceren. alles wat werkt of niet werkt. Elk assay. Elke vraag die ik heb, elk experiment, nee elke gedachte aan een experiment. En ik wil het gewoon opschrijven en online zetten. Zelf.
SLIDE zo ziet het eruit. Ik noem het REBLAB. Die OpenKitchenScience is niet nieuw, er zijn al een aantalw etenschappers die radicaal transparant werken. Op sommige plekken heet dat Open Notebook Science. Een mooi voorbeeld http://labscribbles.com/is waar je het onderzoek kan volgen van dr. Rachel Harding over het Huntingtin eiwit.
Peer review
Laten we het over peer review hebben. Want dat is de belangrijkste wetenschaps-“regel” die ik breek. Ik deel dingen zonder of voorafgaand aan peer review. Nu geloof ik dat peer review een belangrijke maat kan zijn van wetenschappelijke kwaliteit. Tegelijkertijd denk ik dat we er teveel mee bezig zijn. Het begint ons tegen te houden. Ik geloof dat het een belangrijke reden is dat we de minder-shiny positieve verhalen niet met elkaar delen enzoveel van onze kennis, lelijke praktische inzichten en negatieve resultaten, voor altijd in het lab opgesloten blijven.
De vondst dat sommige bacterien alleen D-lactaat maken terwijl andere alleen L-lactaat maken. 100 manieren waarop je niet een reagens in oplossing krijgt. Waarom deelden we dat soort dingen niet? Omdat het publicatieproces, deels dankzij peer review, heel veel tijd, aandacht en energie kost. Niet alleen van jou, maar ook van je collega’s.
Reproduceerbaarheids crisis
Dat is ook de reden dat we, ondanks dat kwaliteitssysteem, ondanks dat we eigenlijk uitsluitend met elkaar communiceren door de peer review filter, de kwaliteit toch te wensen over laat. We maken momenteel een reproduceerbaarheidscrisis door. SLIDE Ongeveer eenderde van de high impact prestigieuze kanker studies waren niet reproduceerbaar. Het was farma die ons aanvankelijk op dat problem wees, “jongens dit zal allemaal wel superfancy wetenschap zijn”. Maar het klopt niet. Het houdt geen stand in onze labs. In de sociale psychologie is het kennelijk erger.
NWO spendeert nu 3 miljoen euro specifiek aan replicatiestudies. Prima initiatief! Maar mijn vraag is: zijn we niet al heel veel bezig met elkaars werk reproduceren? We noemen het alleen anders en we zijn normaal gesproken niet erg blij wanneer het gebeurt. We noemen het “scooping”. In Open Kitchen Science is daarom peer review de zilveren standaard van wetenschapskwaliteit. Gescoopt worden is de gouden standaard. Als je resultaten in een onafhankelijk lab in andere handen stand houden, dan pas vertrouw ik ze.
Hier een aaname: ze wisten allang in het kankerveld dat er verschillende dingen werden gevonden. Het is een ontzettend competitief veld. Er moeten zeker 20 labs zijn die wereldwijd aan dezelfde oncogenen werken en dezelfde mutaties. Misschien, in een wetenschappelijke wereld waar de standard is om alles te delen, niet alleen de positieve uitkomsten maar ook de negatieve insignificante uitkomsten, als de gouden standaard reproductie en niet review is, hadden we het allang geweten. Was ons onderzoek robuuster en nuttiger en relevanter voor de patiënt.
En die onderzoekers, die fraudeerden ook niet. Hun werkelijkheid was alleen waar in het kunstmatige miniwereldje wat ze hadden geschapen. Met hun specifieke weefselkweek, of alleen hun muizen, of alleen hun lab. Problemen die je niet ontdekt met peer review, want de data zien er spectaculair uit. Ik wil dat mijn resultaten robuust en reproduceerbaarheid zijn in zoveel mogelijk kunstmatige minimilieus in labs. En juist wanneer er andere assays worden gebruikt, en andere methodes en net andere stammen, en dezelfde resultaten worden geboekt, dan vertrouw ik ze. Ik wil zo dicht mogelijk bij de echte wereld komen. En op een bepaald moment zal ik peers vragen om er een blik op te werpen, maar nogmaals, zilveren standaard.
Het post-academische aspect
Geen funding hebben. Hobby wetenschap. Vrijwilliger zijn. Het is heerlijk om door niemands hoepels te hoeven springen. Nu moet ik toegeven dat het deels is omdat ik een beetje ongeleid projectiel ben, en niet zo goed tegen autoriteit kan, en heel erg gelukig wordt van onafhankelijkheid, deels omdat het kan omdat ik van het schrijven kan leven, en deels omdat ik niet de papers en de output heb die mijn kansen op een beurs erg groot maken. Maar het ziet er soms ook niet bepaald aantrekkelijk uit, die academische carrière. Ik zie veel teleurstelling, cynisme, negativiteit, mensen die moe zijn van de competitieve en de politieke aspecten en de machtsspelletjes en alles wat er verder bij die prachtige baan komt kijken.
Waar ik denk dat open wetenschap wel de toekomst is, denk ik niet dat dit post-academic wetenschap de toekomst is voor iedereen en dat iedereen dit moet ambieren. Maar ook hier raakt het een belangrijk punt en dat is dat we als wetenschap heel verkwistend zijn met onze wetenschappelijke kwaliteiten. Het is best zeldzaam dat iemand die de academie verlaat nog blijft bijdragen. Is dat niet vreemd? Al die onderzoekskwaliteiten en expertise en kennis waar we publiek geld in hebben geinvesteerd vloeien weg. Waarom maken we ons daar niet meer zorgen over? Zou het niet heel waardevol zijn als we op een of andere manier die mensen betrokken weten te houden?
Wetenschap is in die zin een van de weinige menselijke activiteiten die we alleen op topsport niveau beoefenen. Dat is niet hetzelfde voor muziek of sport of kunst, je kan al die activiteiten ook in een andere hoedanigheid beoefenen dan een professionele betaalde baan. Ik geloof dat wetenschap uiteindelijk dezelfde kansen moet bieden.
SLIDE De praktijk! Eerste stappen richting Open Kitchen Science
SLIDE Open wetenschap voor het Spaarne ziekenhuis?
Het grappige is dat telkens wanneer ik artsen spreek, dat ik het gevoel krijg dat ik helemaal met niets nieuws bezig ben. Het is helemaal niet radicaal wat ik doe. Jullie artsen die zich in STZ ziekenhuizen bezig houden met wetenschap doen dat heel regelmatig onbetaald, naast een meer dan voltijdse baan vol patienten zorg. In avonduren, in vrije tijd.
Bevindingen die vaak ontzettend belangrijk zijn, praktische inzichten, met je neus er boven op, resultaten die vaak veel directer invloed hebben op de kwaliteit van leven en de vooruitzichten van patiënten.
Nog een opmerkelijke constatering. Heel veel artsen zijn tegenwoordig gepromoveerd. Dus het barst hier van de researchskills. Ik weet niet of er een omgeving is waar er buiten de academie en officiele onderzoeksinstituten zoveel PhD’s rondlopen als in een ziekenhuis.
In feite doen jullie ook post-academische wetenschap. Maar ondanks dat wetenschap voor jullie vaak is wat jullie er naast doen, lijkt het erop dat jullie je nog steeds wel laten gijzelen door een enorme hoeveelheid hoepels en de rigide communicatiecultuur van de academische wetenschap.
En eigenlijk is dat bizar, want ik vroeg het net even hier maar jullie hebben volgens mij ook geen beste toegang tot de literatuur. De onderzoekers die ik sprak zeiden, oh nee, dat regel ik via iemand met een account. Gekke vraag misschien, maar als je zelf moet bedelen en je in bochten moet wringen om nota bene je eigen papers te kunnen lezen, gewoon omdat de abonnementen belachelijk duur zijn voor een ziekenhuis, waarom zou je dan nog bij dat soort uitgevers publiceren. Waarom zou je dan nog gratis content creëren? Waarom zou je dan in je vrije tijd tekstjes typen voor uitgeverijen die daarna te duur zijn om terug te kopen?
Wat dat betreft liggen hier juist kansen voor open wetenschap. juist wanneer die papers niet per se meer nodig zijn voor je salaris, voor je carriere, of om je volgende grantaanvraag geaccepteerd te krijgen. Juist als die papers niet meer je belangrijkste output zijn, maar liefde werk zijn, gedreven door idealisme, kennis doorgeven en verspreiden, drive om je horizon te verbreden en de kennis in je vakgebied te vergroten.
Ik hoop dat ik jullie heb kunnen inspireren om open science te omarmen. Je hoeft het niet zo radicaal aan te pakken als ik. Je kunt gewoon eens heel voorzichtig. Een poster online zetten. Een praatje op een congres van te voren opnemen en online zetten. Een commentaar op een paper op een blog plaatsen. Een case report niet naar een officiele reviewer sturen maar gewoon eens online zetten. Sociale media kan je gebruiken om je publicaties te delen.
Het zou juist weleens hier in het ziekenhuis kunnen zijn, buiten de hotshot science van de academische ziekenhuizen waar je ermee kan beginnen. Ik hoop van harte dat het jullie lukt.
Two hypotheses on bacterial metabolism in the vagina
Hello everyone, first bigger update of the project since December. Things have been progressing slowly but surely. This will be a long blog post so I’ll start with an overview.
Overview
1 During my talk in Groningen at the Young Academy I had the pleasure to meet dr. Alicia Brandt who aspires to remain connected and contributing to science even though she left her official scientific position. Her expertise is bacterial glycobiology and she performed a “dbCan” survey to find all carbohydrate-active enzymes (CAZymes) in the genomes of the different Lactobacillus crispatus strains that were isolated from BV+ and BV- women.
2 She also presented a intriguing hypothesis on possible sugar sources in the vaginal environment. Although I am looking at glycogen as the main vaginal sugar source for acidification by lactobacilli, a different sugar called “levan” could also function as an interesting intermediate metabolite for lactic acid production. I include Alicia’s hypothesis, her analysis on the presence of levansucrases encoding genes in the human microbiome and insights into possible L. crispatus levan metabolism.
3 One of the genes that may be involved in glycogen metabolism is a putative extracellular S-layer linked type 1 pullulanase. Zooming in on this gene I several variants present in the different L. crispatus genomes. Is this a possible match with glycogen metabolic abilities? Nothing final yet, but a promising lead to look further into.
Alicia Brandt: dbCan analysis and glycogen-active enzymes
Since I started with REBLAB I encountered several people who share their own stories about their departure from science and how this affects them. Sometimes these people find the time and energy to still contribute to science besides their regular jobs. One of these people is Alicia Brandt (previously Alicia Lammerts van Bueren as she is known in the glycobiology field) who left her job as a postdoc last year for a job in a supporting role at the University of Groningen. She expressed the strong wish to keep being involved and to share her knowledge and skill. It’s a happy coincidence that her expertise is exactly what I am looking for: glycobiology and even glycogen metabolism of bacteria. She offered to help me out with several analyses. First thing she did was to perform an analysis using the so-called dbcan server, detecting the presence of several glycoproteins that potentially be involved in glycogen metabolism.
-add: operon
-various glycogen active enzymes
-N-terminus gh13_13
Levansucrase and the vaginal microbiota
Alicia also brought up a second possible sugar source that lactobacilli could use to produce lactic acid and acidify the vagina: levan. From her analysis it seems that especially the bacterial communities colonizing the mouth and vagina are equipped to synthesize levan. The following text is all Alicia’s. Please find the protocol she used here and the excel sheet with complete overview of the dbcan results here.
Bacterial levansucrase in the vagina
Levansucrases (EC2.4.1.10) are enzymes that catalyse the transfructosylation of sucrose into the polysaccharide levan (ref, ref). These enzymes are found within the CAZy family GH68. GH68 enzymes use sucrose as their preferential donor substrate. Many of the enzymes can create very long levan-type fructans (catalyzed by levansucrases) or inulin-type of fructans (catalyzed by inulosucrases), as well as fructooligosacharides (FOS). However, some GH68 enzymes can also use fructan as donor substrate (in the absence of sucrose or at a high fructan/sucrose ratio) (see CAZypedia).
Levans and inulins are synthesized by bacteria and some plant species (for example chickory root inulin). In lactic acid bacteria, levan is more commonly produced over inulin. In these lactic acid bacteria, levan and inulin act as exopolysaccharides that provide a protective layer around the bacteria, shielding it from the stresses of the external environment (ref). Levans can range in size from 6-kestose (DP3) up to MDa in size. It is a biologically important compound and is used in manufacturing processes as thickening agents. It was demonstrated to be a beneficial prebiotic source. Levan and inulin are degraded via the action of GH32 enzymes (CAZypedia link), thus bacteria that encode this family of enzyme are typically capable of using levan as a nutritional source. Levans can range in size from 6-kestose (DP3) up to MDa in size (for example 130MDa or 1.3 x 108 Da in Brenneria goodwinii). Levan and inulin are degraded via the action of GH32 enzymes (CAZypedia link), thus bacteria that encode this family of enzyme are typically capable of using levan or inulin as a nutritional source.
Levan and inulin are biologically important compounds and are used in manufacturing processes as thickening agents. Levan has been demonstrated to be beneficial prebiotic source.
Fig 2: Synthesis of levan by GH68 enzymes (image from here)

Glucansucrases (EC2.4.1.5) (family GH70, see CAZypedia) are processive enzymes similar to levansucrases. However these enzymes carry out transglucosylation of sucrose into alpha-glucan polymers, such as dextran, reuteran, alternan (ref). These enzymes are also mainly encoded by lactic acid bacteria and are used by these bacteria for the production of alpha-glucan exopolysaccharides. The enzymes and exopolysaccharide products also have great interest to industry in food and nutrition applications.
Fig 3: Levan versus alpha-glucan synthesis by fructansucrases and glucansucrases levan/inulin alpha-glucan
Levansucrases in the microbiome
Lactic acid bacteria make up a significant portion of our human, mainly gut, microbiota. The exopolysaccharides they produce using glucan and levan sucrases are biologically important molecules. The best known example are the exopolysaccharides produced by the oral bacteria Streptococcus mutans. In our mouths, S. mutans takes dietary sucrose and produces a sticky biofilm for attachment to the surface of tooth enamel which is the most significant contributor to tooth decay and dental caries. Glucan and levan exopolysaccharide have also been shown to serve as important nutritional sources for strains of bacteria that reside in our gastrointestinal tracts (exopolysaccharides as microbial products have prebiotic effects, my research in Groningen). Therefore, I was interested to find out in what scale the presence of genes encoding levansucrases and glucansucrases are in metagenomic sequences of human microbiome samples.
In order to determine the abundance of GH68 and GH70 enzymes in human microbiome metagenomic data, I performed a metagenomic search in Human Microbiome Project database from the JGI-DOE Integrated Microbial Genomes & Microbiomes(IMG/M) system (This analysis is from August 2014, results may be drastically different now with so much more metagenomic sequencing data available). Within the database I performed a “Find Function – Enzymes” search using EC numbers EC2.4.1.10 (levansucrases) and EC2.4.1.5 (glucansucrases). Within the database, I selected to search within metagenomic data from microbiome sample sites including the digestive system (oral, large intestine and small intestine) stool, and reproductive system. All results were tabulated in excel file (see folder).
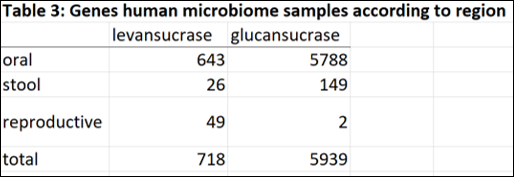
The majority of genes encoding glucansucrase and levansucrase enzymes are found amongst the microbiota of the oral cavity. In the mouth and stool, glucan sucrases are far more abundant than levansucrases. However in the human vagina, where bacterial levels are much lower and less diverse than mouth and gut, levansucrases are more abundant than glucansucrases (49 versus 2). Thus the questions raised are:
1. Why are levansucrases more abundant than glucansucrases in the human vagina?
2. What is the importance of levan in the vagina?
3. Is levan metabolized by other species in the human vagina?
4. Is levan biologically important to the human vagina? (Health versus disease state?)
5. What is the source of the donor substrate (sucrose or fructan) for fructansucrases/levansucrases in the human vagina?
The main carbohydrate sources that would be found in the human vagina are glycogen, human N and O linked glycans, and microbially synthesized glycans, such as exopolysaccharides. Other metabolites include lactate, succinate, polyamines, amino acid breakdown products, dipeptides, These metabolites strongly vary depending on the microbiome. However, none of these sources would contribute to levan synthesis and provide the essential sucrose which acts as a donor and acceptor substrate. Where could alternative substrates for levan synthesis be found in the vagina?
In sexually active women, a source of for the sucrose or fructan donor substrate for levan synthesis could be found in semen. The limited analysis of the carbohydrate content of human semen carried out in the 1940’s and 1960’s has identified that fructose is abundant from 1- 5 mg/ml.
(there are only some reports from the 40’s and 60’s but, funny enough, there are no recent reports of the carbohydrate analysis of human semen. This should be done because our modern methods could really identify the carbohydrate composition of human semen.)
Hypothesis:
Levan is a biologically important polysaccharide in the human vagina.
Hypothesis 1: The source of the donor substrate for fructansucrase activity (sucrose/fructose/fructans) comes from human semen.
Hypothesis 2: lactic acid bacteria harboring fructan/levansucrase enzymes synthesize levan from the fructose/fructans found in male semen. Since the presence of semen is transient, and carbohydrate sources in the human vagina are limited, levan synthesis by lactic acid bacteria strains in the vagina serves three functions
-At times when fructose/fructans are abundant (such as after sexual intercourse), lactic acid bacteria synthesize levan in order to create an equilibrium in their environment.
-At times when carbohydrate sources are limited, levan can be used as a nutritional source and metabolized by bacteria in the human vagina (that contain GH32 enzymes).
-Metabolism of levan by bacteria in the human vagina contribute to the acidification of the vaginal environment, which contribute to vaginal health.